

Differences between human slow N-acetyltransferase 2 alleles in levels of 4-aminobiphenyl-induced DNA adducts and mutations
- PMID: 19682468
- PMCID: PMC2783811
- DOI: 10.1016/j.mrfmmm.2009.08.003
Differences between human slow N-acetyltransferase 2 alleles in levels of 4-aminobiphenyl-induced DNA adducts and mutations
Abstract
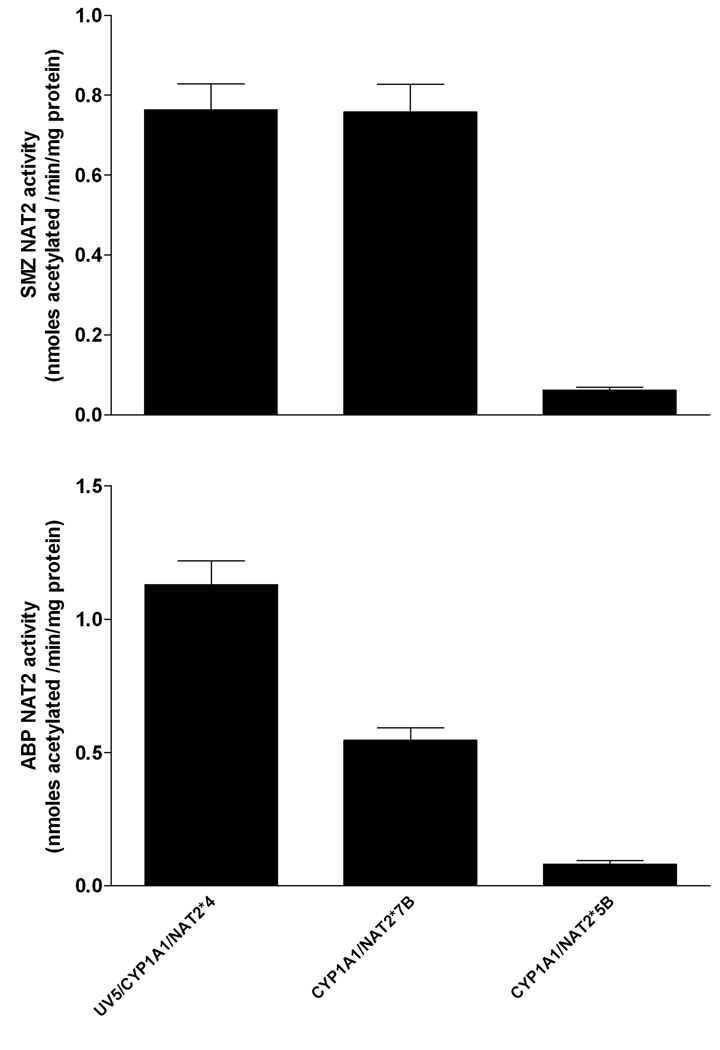
Aromatic amines such as 4-aminobiphenyl (ABP) require biotransformation to exert their carcinogenic effects. Genetic polymorphisms in biotransformation enzymes such as N-acetyltransferase 2 (NAT2) may modify cancer risk following exposure. Nucleotide excision repair-deficient Chinese hamster ovary (CHO) cells stably transfected with human cytochrome P4501A1 (CYP1A1) and a single copy of either NAT2*4 (rapid acetylator), NAT2*5B (common Caucasian slow acetylator), or NAT2*7B (common Asian slow acetylator) alleles (haplotypes) were treated with ABP to test the effect of NAT2 polymorphisms on DNA adduct formation and mutagenesis. ABP N-acetyltransferase catalytic activities were detectable only in cell lines transfected with NAT2 and were highest in cells transfected with NAT2*4, lower in cells transfected with NAT2*7B, and lowest in cells transfected with NAT2*5B. Following ABP treatment, N-(deoxyguanosin-8-yl)-4-aminobiphenyl (dG-C8-ABP) was the primary adduct formed. Cells transfected with both CYP1A1 and NAT2*4 showed the highest concentration-dependent cytotoxicity, hypoxanthine phosphoribosyl transferase (hprt) mutants, and dG-C8-ABP adducts. Cells transfected with CYP1A1 and NAT2*7B showed lower levels of cytotoxicity, hprt mutagenesis, and dG-C8-ABP adducts. Cells transfected with CYP1A1 only or cells transfected with both CYP1A1 and NAT2*5B did not induce cytotoxicity, hprt mutagenesis or dG-C8-ABP adducts. ABP-DNA adduct levels correlated very highly (r>0.96) with ABP-induced hprt mutant levels following each treatment. The results of the present study suggest that investigations of NAT2 genotype or phenotype associations with disease or toxicity could be more precise and reproducible if heterogeneity within the "slow" NAT2 acetylator phenotype is considered and incorporated into the study design.
Figures





References
-
- Stabbert R, Schafer KH, Biefel C, Rustemeier K. Analysis of aromatic amines in cigarette smoke. Rapid Commun Mass Spectrom. 2003;17:2125–2132. - PubMed
-
- Baan R, Straif K, Grosse Y, Secretan B, Ghissassi FEl, Bouvard V, Benbrahim-Tallaa L, Cogliano V. Carcinogenicity of some aromatic amines, dyes, organic dyes, and related exposures. Lancet Oncol. 2008;9:322–323. - PubMed
-
- Kim D, Guengerich FP. Cytochrome P450 activation of arylamines and heterocyclic amines. Annu Rev Pharmacol Toxicol. 2005;45:27–49. - PubMed
-
- Dorrenhaus, Muller T, Roos PH. Increased CYP1A1 expression in human exfoliated urothelial cells of cigarette smokers compared to non-smokers. Arch Toxicol. 2007;81:19–25. - PubMed
-
- Tsuneoka Y, Dalton TP, Miller ML, Clay CD, Shertzer HG, Talaska G, Medvedovic M, Nebert DW. 4-aminobiphenyl-induced liver and urinary bladder DNA adduct formation in Cyp1a2(−/−) and Cyp1a2(+/+) mice. J Natl Cancer Inst. 2003;95:1227–1237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
