

Influence of a model human defensive peroxidase system on oral streptococcal antagonism
- PMID: 19684069
- PMCID: PMC2888128
- DOI: 10.1099/mic.0.031310-0
Influence of a model human defensive peroxidase system on oral streptococcal antagonism
Abstract
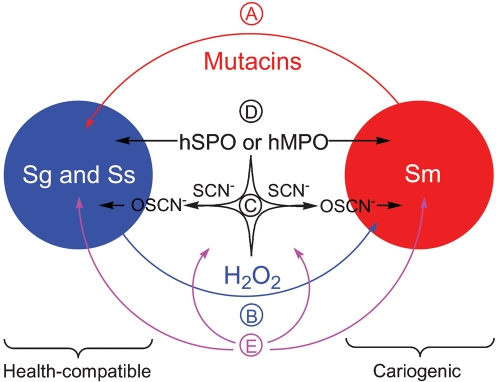
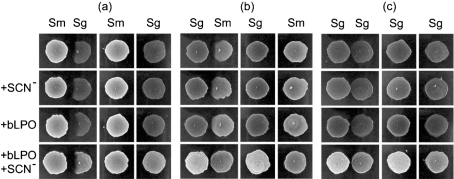
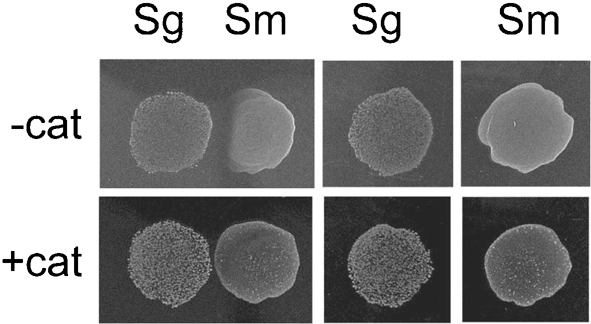
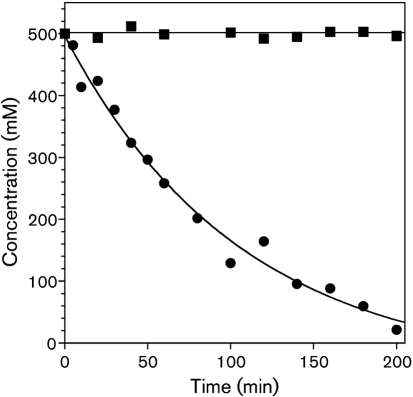
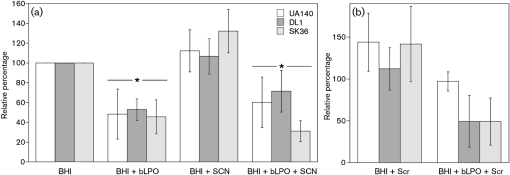
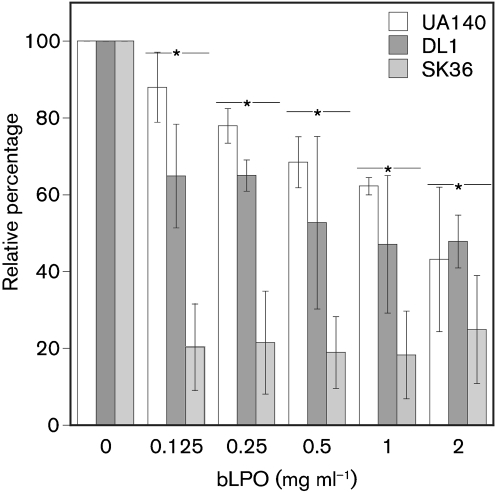
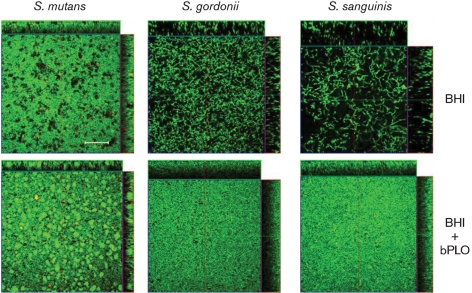
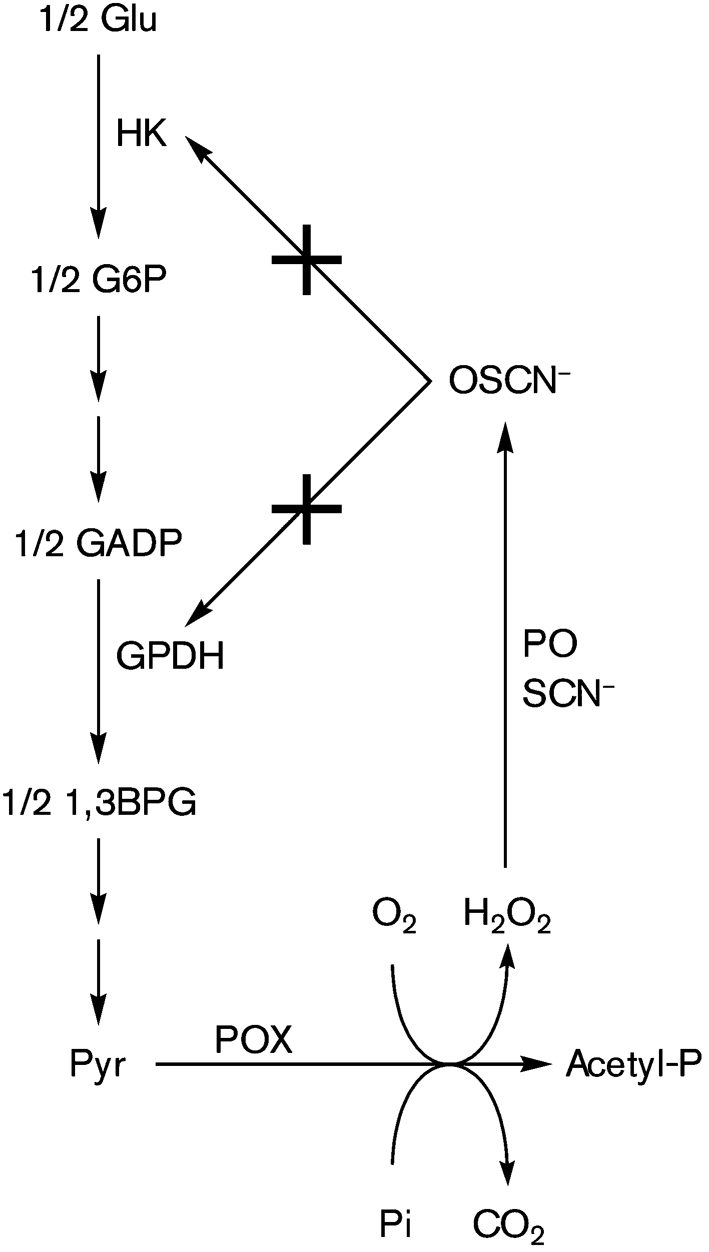
Streptococcus is a dominant genus in the human oral cavity, making up about 20 % of the more than 800 species of bacteria that have been identified, and about 80 % of the early biofilm colonizers. Oral streptococci include both health-compatible (e.g. Streptococcus gordonii and Streptococcus sanguinis) and pathogenic strains (e.g. the cariogenic Streptococcus mutans). Because the streptococci have similar metabolic requirements, they have developed defence strategies that lead to antagonism (also known as bacterial interference). S. mutans expresses bacteriocins that are cytotoxic toward S. gordonii and S. sanguinis, whereas S. gordonii and S. sanguinis differentially produce H(2)O(2) (under aerobic growth conditions), which is relatively toxic toward S. mutans. Superimposed on the inter-bacterial combat are the effects of the host defensive mechanisms. We report here on the multifarious effects of bovine lactoperoxidase (bLPO) on the antagonism between S. gordonii and S. sanguinis versus S. mutans. Some of the effects are apparently counterproductive with respect to maintaining a health-compatible population of streptococci. For example, the bLPO system (comprised of bLPO+SCN(-)+H(2)O(2)) destroys H(2)O(2), thereby abolishing the ability of S. gordonii and S. sanguinis to inhibit the growth of S. mutans. Furthermore, bLPO protein (with or without its substrate) inhibits bacterial growth in a biofilm assay, but sucrose negates the inhibitory effects of the bLPO protein, thereby facilitating adherence of S. mutans in lieu of S. gordonii and S. sanguinis. Our findings may be relevant to environmental pressures that select early supragingival colonizers.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources