

Origins of correlated activity in an olfactory circuit
- PMID: 19684589
- PMCID: PMC2751859
- DOI: 10.1038/nn.2376
Origins of correlated activity in an olfactory circuit
Abstract
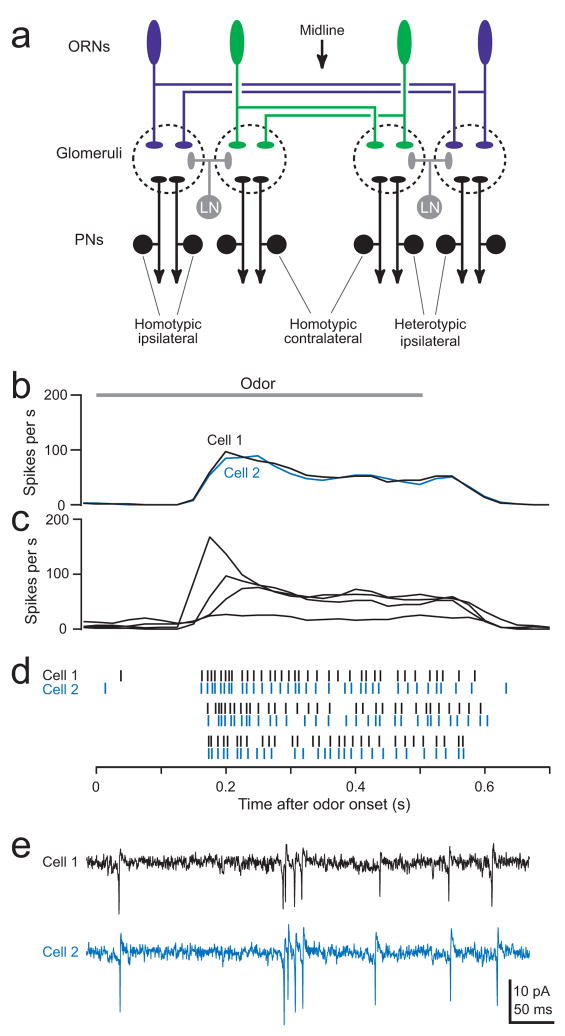
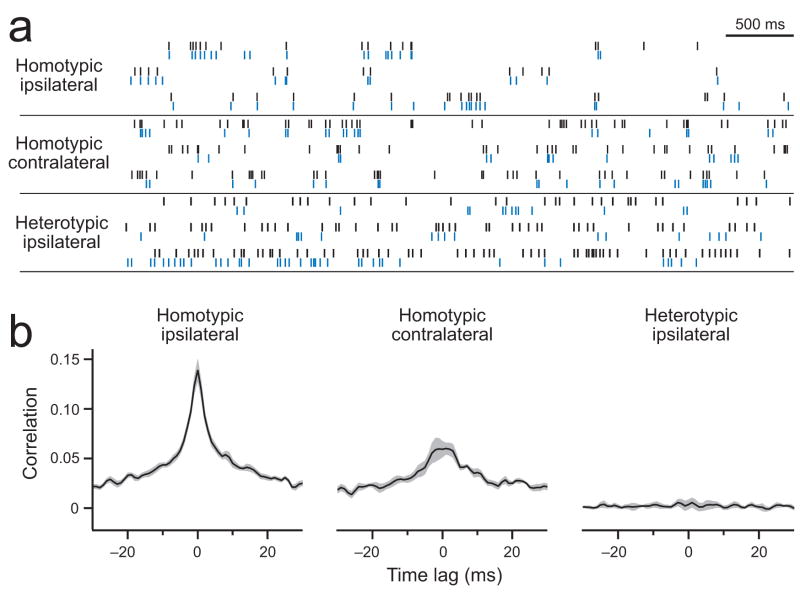
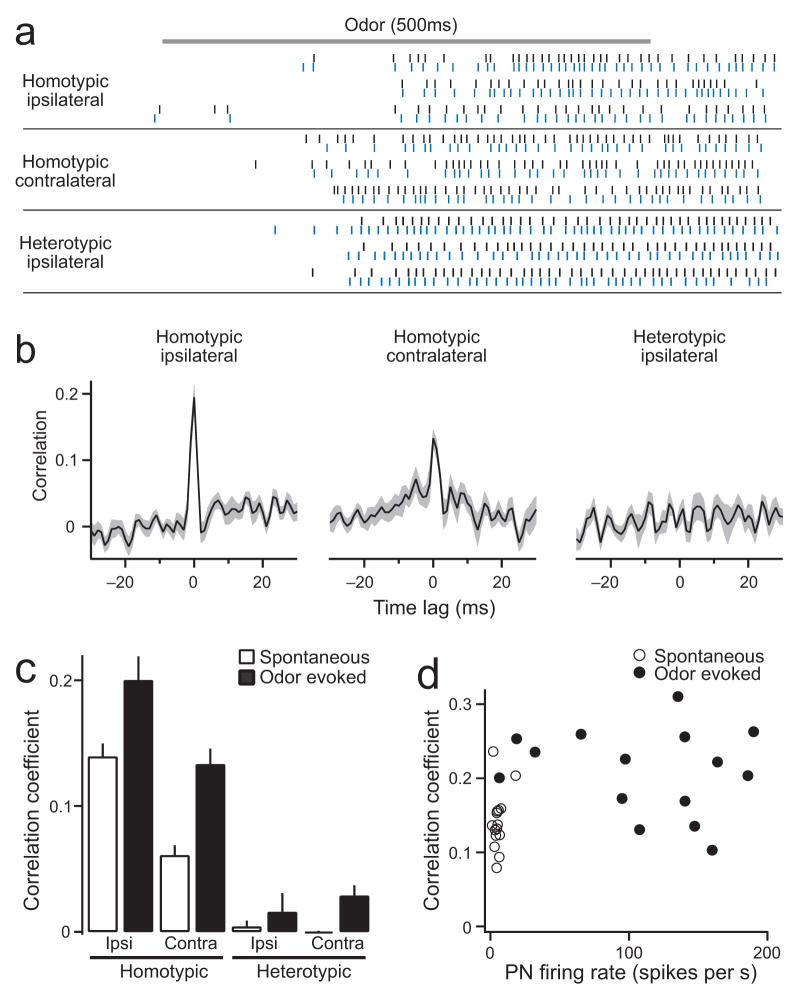
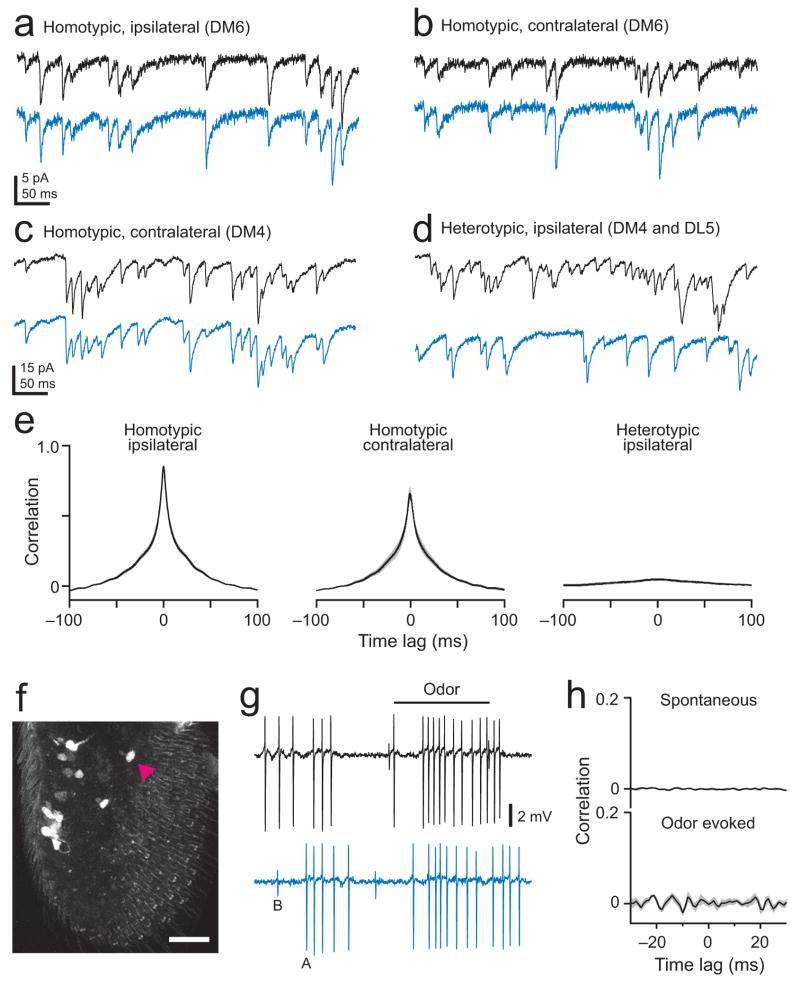
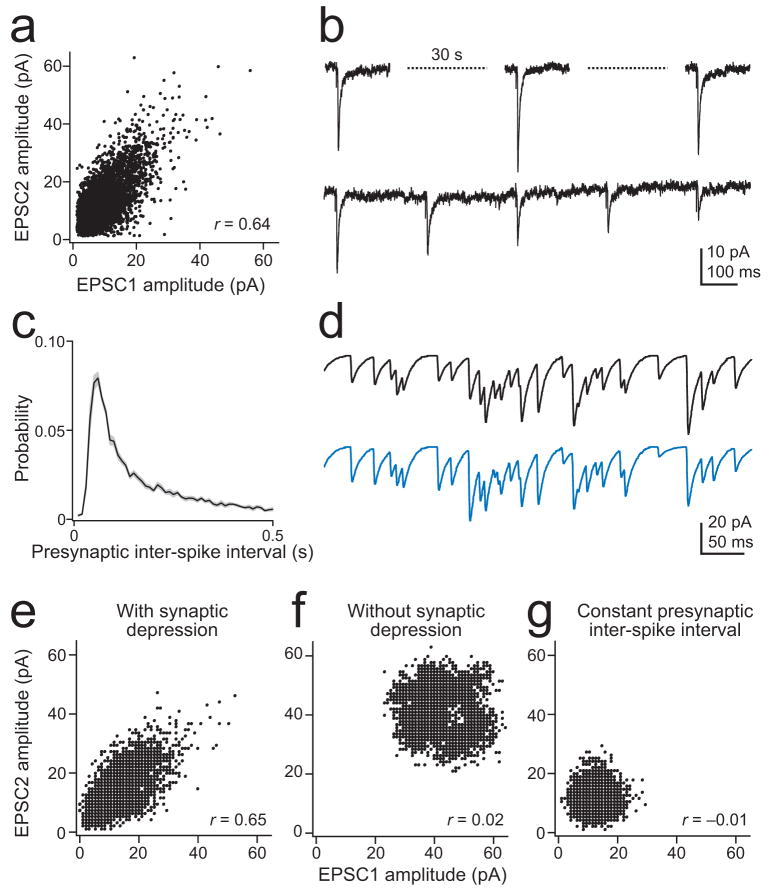
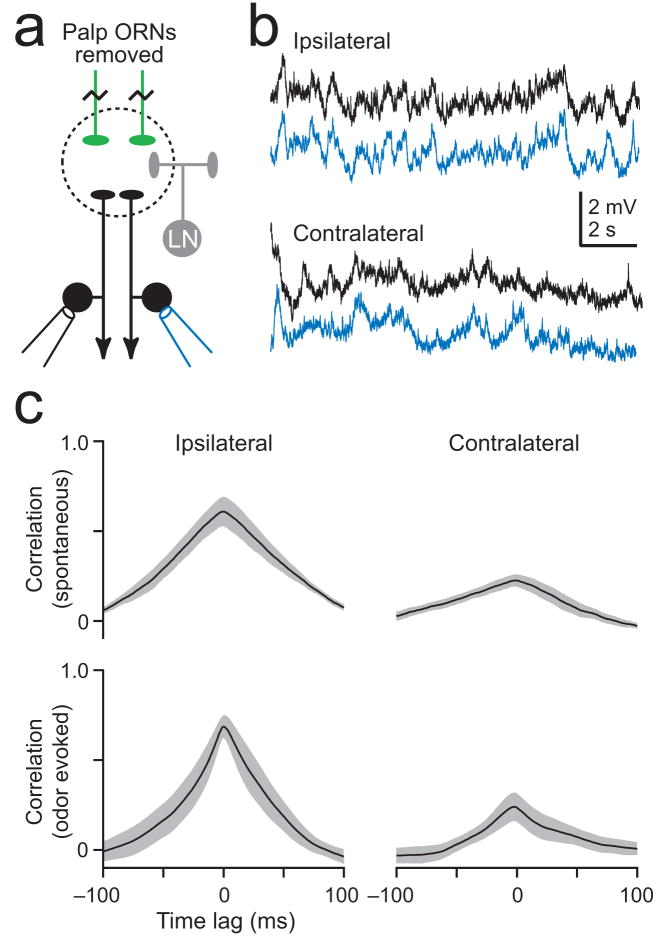
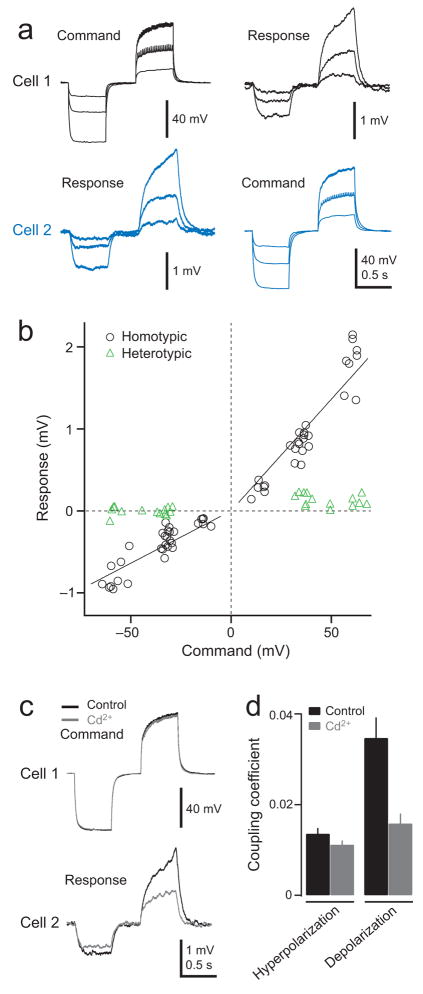
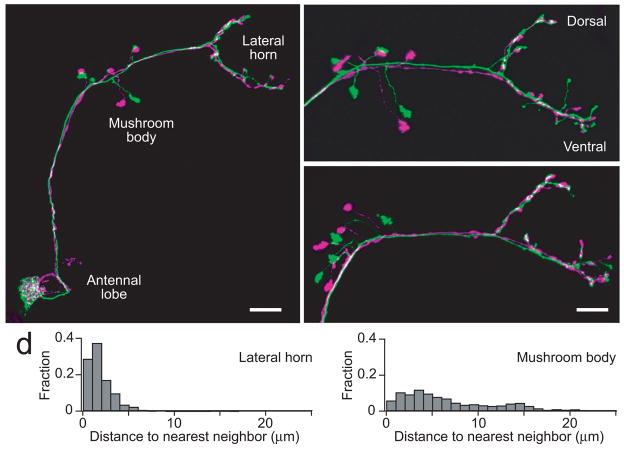
Multineuronal recordings often reveal synchronized spikes in different neurons. The manner in which correlated spike timing affects neural codes depends on the statistics of correlations, which in turn reflects the connectivity that gives rise to correlations. However, determining the connectivity of neurons recorded in vivo can be difficult. We investigated the origins of correlated activity in genetically labeled neurons of the Drosophila antennal lobe. Dual recordings showed synchronized spontaneous spikes in projection neurons (PNs) postsynaptic to the same type of olfactory receptor neuron (ORN). Odors increased these correlations. The primary origin of correlations lies in the divergence of each ORN onto every PN in its glomerulus. Reciprocal PN-PN connections make a smaller contribution to correlations and PN spike trains in different glomeruli were only weakly correlated. PN axons from the same glomerulus reconverge in the lateral horn, where pooling redundant signals may allow lateral horn neurons to average out noise that arises independently in these PNs.
Figures
References
-
- Usrey WM, Reid RC. Synchronous activity in the visual system. Annu Rev Physiol. 1999;61:435–56. - PubMed
-
- Averbeck BB, Latham PE, Pouget A. Neural correlations, population coding and computation. Nat Rev Neurosci. 2006;7:358–66. - PubMed
-
- Meister M, Lagnado L, Baylor DA. Concerted signaling by retinal ganglion cells. Science. 1995;270:1207–10. - PubMed
-
- Dan Y, Alonso JM, Usrey WM, Reid RC. Coding of visual information by precisely correlated spikes in the lateral geniculate nucleus. Nat Neurosci. 1998;1:501–7. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
