

Microbial communication, cooperation and cheating: quorum sensing drives the evolution of cooperation in bacteria
- PMID: 19684853
- PMCID: PMC2722019
- DOI: 10.1371/journal.pone.0006655
Microbial communication, cooperation and cheating: quorum sensing drives the evolution of cooperation in bacteria
Abstract
An increasing body of empirical evidence suggests that cooperation among clone-mates is common in bacteria. Bacterial cooperation may take the form of the excretion of "public goods": exoproducts such as virulence factors, exoenzymes or components of the matrix in biofilms, to yield significant benefit for individuals joining in the common effort of producing them. Supposedly in order to spare unnecessary costs when the population is too sparse to supply the sufficient exoproduct level, many bacteria have evolved a simple chemical communication system called quorum sensing (QS), to "measure" the population density of clone-mates in their close neighborhood. Cooperation genes are expressed only above a threshold rate of QS signal molecule re-capture, i.e., above the local quorum of cooperators. The cooperative population is exposed to exploitation by cheaters, i.e., mutants who contribute less or nil to the effort but fully enjoy the benefits of cooperation. The communication system is also vulnerable to a different type of cheaters ("Liars") who may produce the QS signal but not the exoproduct, thus ruining the reliability of the signal. Since there is no reason to assume that such cheaters cannot evolve and invade the populations of honestly signaling cooperators, the empirical fact of the existence of both bacterial cooperation and the associated QS communication system seems puzzling. Using a stochastic cellular automaton approach and allowing mutations in an initially non-cooperating, non-communicating strain we show that both cooperation and the associated communication system can evolve, spread and remain persistent. The QS genes help cooperative behavior to invade the population, and vice versa; cooperation and communication might have evolved synergistically in bacteria. Moreover, in good agreement with the empirical data recently available, this synergism opens up a remarkably rich repertoire of social interactions in which cheating and exploitation are commonplace.
Conflict of interest statement
Figures
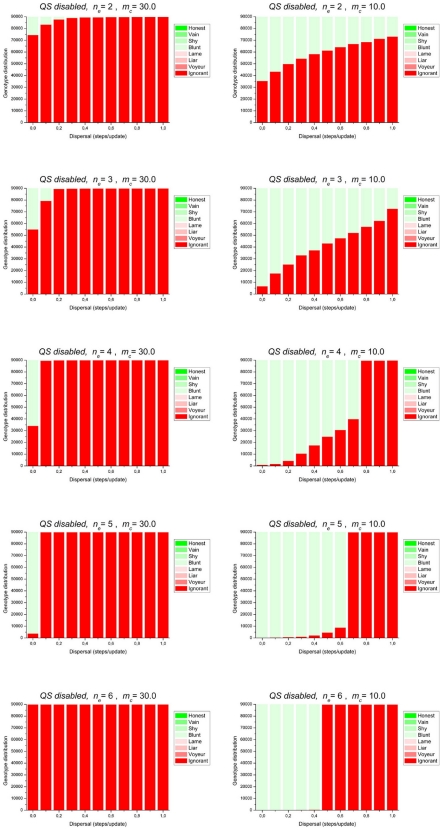
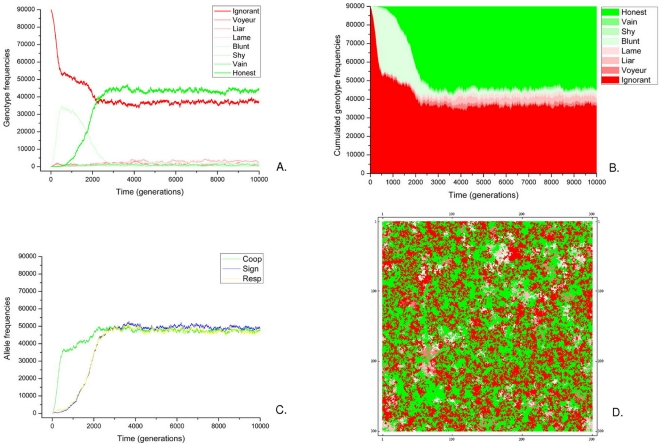
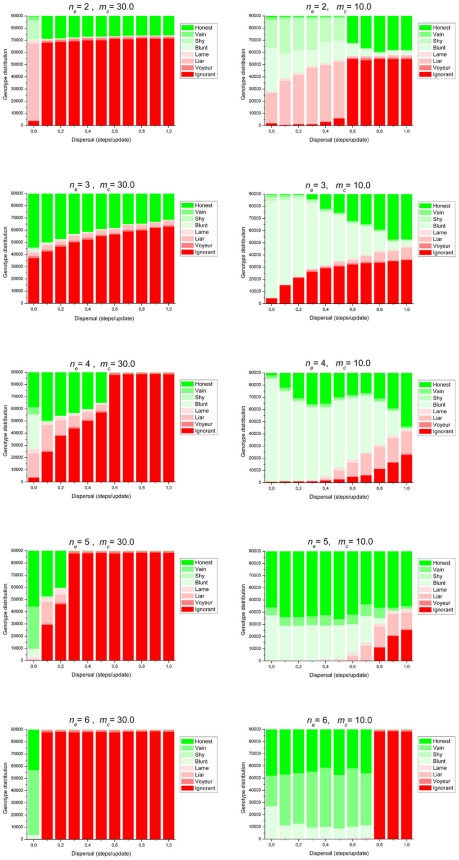
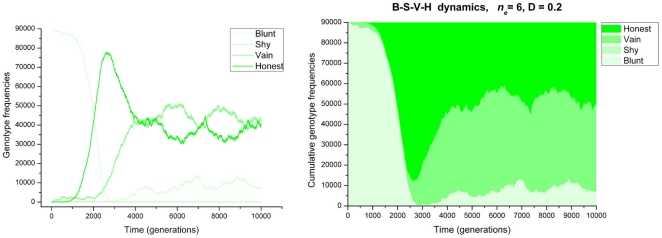
References
-
- Hardin G. The tragedy of the commons. Science. 1968;162:1243–1244. - PubMed
-
- Hamilton WD. The genetical evolution of social behaviour, I & II. J Theor Biol. 1964;7:1–52. - PubMed
-
- West SA, Griffin AS, Gardner A. Evolutionary explanations for cooperation. Curr Biol. 2007;17:R661–72. - PubMed
-
- Queller DC. Quantitative genetics, inclusive fitness, and group selection. American Naturalist. 1992;139:540–558.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
