

Functional characterization of the interactions between endosomal adaptor protein APPL1 and the NuRD co-repressor complex
- PMID: 19686092
- PMCID: PMC2762692
- DOI: 10.1042/BJ20090086
Functional characterization of the interactions between endosomal adaptor protein APPL1 and the NuRD co-repressor complex
Abstract
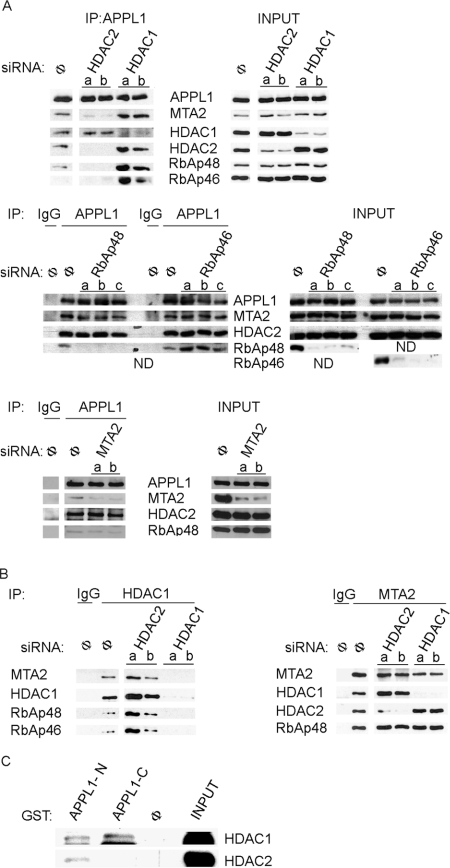
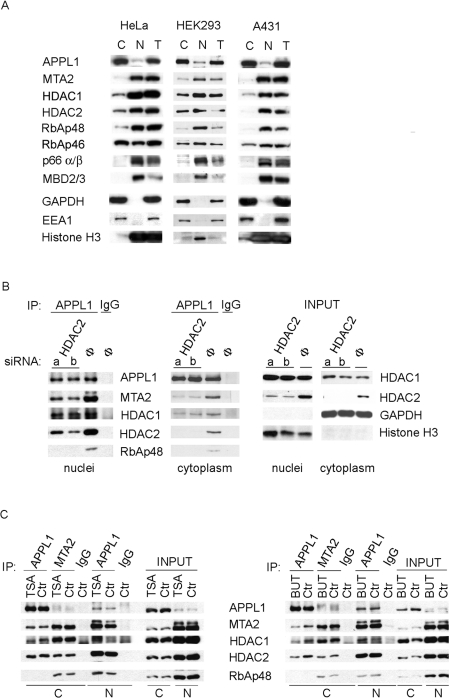
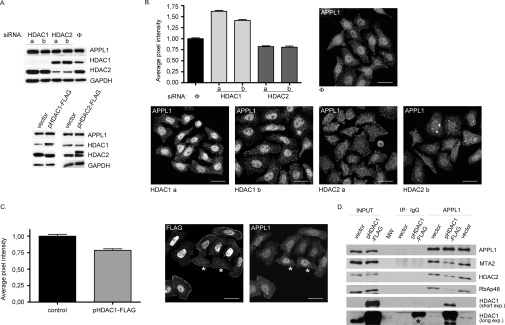
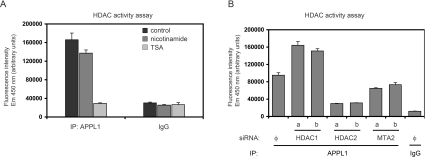
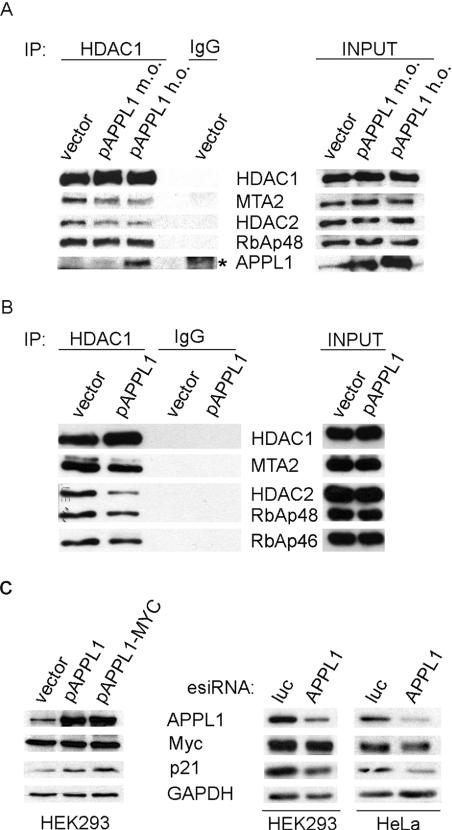
Multifunctional adaptor protein APPL1 [adaptor protein containing PH (pleckstrin homology) domain, PTB (phosphotyrosine binding) domain and leucine zipper motif] belongs to a growing group of endocytic proteins which actively participate in various stages of signalling pathways. Owing to its interaction with the small GTPase Rab5, APPL1 localizes predominantly to a subpopulation of early endosomes but is also capable of nucleocytoplasmic shuttling. Among its various binding partners, APPL1 was reported to associate with the nuclear co-repressor complex NuRD (nucleosome remodelling and deacetylase), containing both nucleosome remodelling and HDAC (histone deacetylase) activities, but the biochemical basis or functional relevance of this interaction remained unknown. Here we characterized the binding between APPL1 and NuRD in more detail, identifying HDAC2 as the key NuRD subunit responsible for this association. APPL1 interacts with the NuRD complex containing enzymatically active HDAC2 but not HDAC1 as the only deacetylase. However, the cellular levels of HDAC1 can regulate the extent of APPL1-NuRD interactions, which in turn modulates the nucleocytoplasmic distribution of APPL1. Increased binding of APPL1 to NuRD upon silencing of HDAC1 promotes the nuclear localization of APPL1, whereas HDAC1 overexpression exerts an opposite effect. Moreover, we also uncovered a NuRD-independent interaction of APPL1 with HDAC1. APPL1 overexpression affects the composition of the HDAC1-containing NuRD complex and the expression of HDAC1 target p21WAF1/CIP1. Cumulatively, these data reveal a surprising complexity of APPL1 interactions with HDACs, with functional consequences for the modulation of gene expression. In a broader sense, these results contribute to an emerging theme of endocytic proteins playing alternative roles in the cell nucleus.
Figures
References
-
- Hoeller D., Volarevic S., Dikic I. Compartmentalization of growth factor receptor signalling. Curr. Opin. Cell Biol. 2005;17:107–111. - PubMed
-
- Le Roy C., Wrana J. L. Clathrin- and non-clathrin-mediated endocytic regulation of cell signalling. Nat. Rev. Mol. Cell. Biol. 2005;6:112–126. - PubMed
-
- Miaczynska M., Pelkmans L., Zerial M. Not just a sink: endosomes in control of signal transduction. Curr. Opin. Cell Biol. 2004;16:400–406. - PubMed
-
- Polo S., Di Fiore P. P. Endocytosis conducts the cell signaling orchestra. Cell. 2006;124:897–900. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous