

Small G proteins in peroxisome biogenesis: the potential involvement of ADP-ribosylation factor 6
- PMID: 19686593
- PMCID: PMC3224584
- DOI: 10.1186/1471-2121-10-58
Small G proteins in peroxisome biogenesis: the potential involvement of ADP-ribosylation factor 6
Abstract
Background: Peroxisomes execute diverse and vital functions in virtually every eukaryote. New peroxisomes form by budding from pre-existing organelles or de novo by vesiculation of the ER. It has been suggested that ADP-ribosylation factors and COPI coatomer complexes are involved in these processes.
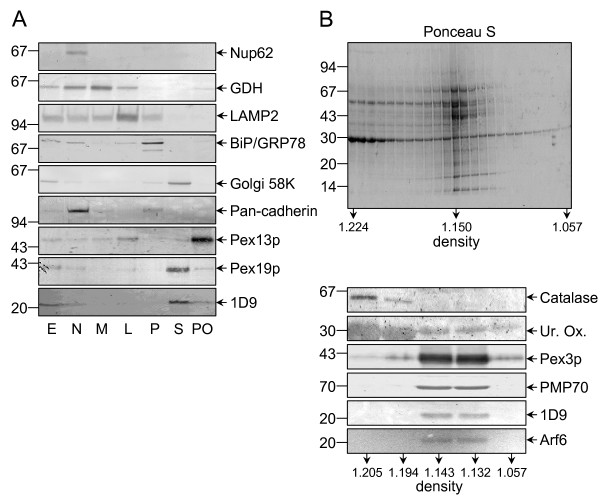
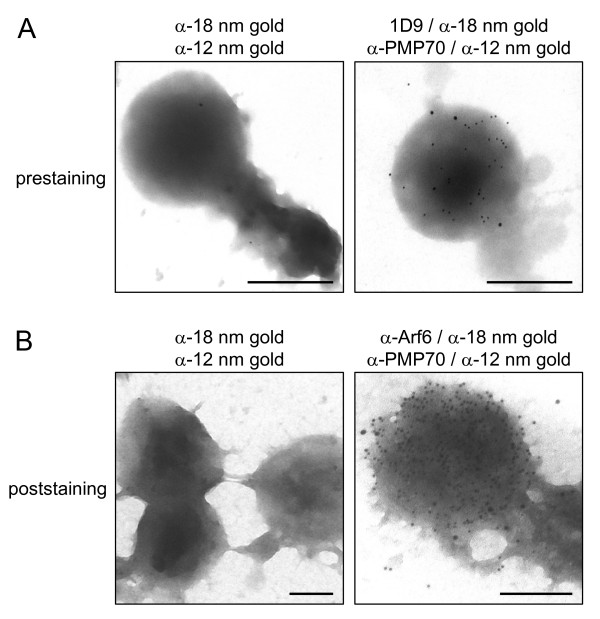
Results: Here we show that all viable Saccharomyces cerevisiae strains deficient in one of the small GTPases which have an important role in the regulation of vesicular transport contain functional peroxisomes, and that the number of these organelles in oleate-grown cells is significantly upregulated in the arf1 and arf3 null strains compared to the wild-type strain. In addition, we provide evidence that a portion of endogenous Arf6, the mammalian orthologue of yeast Arf3, is associated with the cytoplasmic face of rat liver peroxisomes. Despite this, ablation of Arf6 did neither influence the regulation of peroxisome abundance nor affect the localization of peroxisomal proteins in cultured fetal hepatocytes. However, co-overexpression of wild-type, GTP hydrolysis-defective or (dominant-negative) GTP binding-defective forms of Arf1 and Arf6 caused mislocalization of newly-synthesized peroxisomal proteins and resulted in an alteration of peroxisome morphology.
Conclusion: These observations suggest that Arf6 is a key player in mammalian peroxisome biogenesis. In addition, they also lend strong support to and extend the concept that specific Arf isoform pairs may act in tandem to regulate exclusive trafficking pathways.
Figures





Similar articles
-
Binding and functions of ADP-ribosylation factor on mammalian and yeast peroxisomes.J Biol Chem. 2005 Oct 14;280(41):34489-99. doi: 10.1074/jbc.M503497200. Epub 2005 Aug 12. J Biol Chem. 2005. PMID: 16100119
-
Small GTPases in peroxisome dynamics.Biochim Biophys Acta. 2016 May;1863(5):1006-13. doi: 10.1016/j.bbamcr.2016.01.004. Epub 2016 Jan 8. Biochim Biophys Acta. 2016. PMID: 26775587 Review.
-
Peroxisome biogenesis: where Arf and coatomer might be involved.Biochim Biophys Acta. 2006 Dec;1763(12):1678-87. doi: 10.1016/j.bbamcr.2006.08.036. Epub 2006 Aug 30. Biochim Biophys Acta. 2006. PMID: 17023067 Review.
-
YHR150w and YDR479c encode peroxisomal integral membrane proteins involved in the regulation of peroxisome number, size, and distribution in Saccharomyces cerevisiae.J Cell Biol. 2003 Apr 28;161(2):321-32. doi: 10.1083/jcb.200210130. Epub 2003 Apr 21. J Cell Biol. 2003. PMID: 12707309 Free PMC article.
-
ARF- and coatomer-mediated peroxisomal vesiculation.Cell Biochem Biophys. 2000;32 Spring:27-36. doi: 10.1385/cbb:32:1-3:27. Cell Biochem Biophys. 2000. PMID: 11330057 Review.
Cited by
-
Human sperm tail proteome suggests new endogenous metabolic pathways.Mol Cell Proteomics. 2013 Feb;12(2):330-42. doi: 10.1074/mcp.M112.020552. Epub 2012 Nov 15. Mol Cell Proteomics. 2013. PMID: 23161514 Free PMC article.
-
Proteomic analysis reveals that the Rab GTPase RabE1c is involved in the degradation of the peroxisomal protein receptor PEX7 (peroxin 7).J Biol Chem. 2013 Feb 22;288(8):6014-23. doi: 10.1074/jbc.M112.438143. Epub 2013 Jan 7. J Biol Chem. 2013. PMID: 23297417 Free PMC article.
-
Lipid droplets and peroxisomes: key players in cellular lipid homeostasis or a matter of fat--store 'em up or burn 'em down.Genetics. 2013 Jan;193(1):1-50. doi: 10.1534/genetics.112.143362. Genetics. 2013. PMID: 23275493 Free PMC article. Review.
-
Peroxisome biogenesis, membrane contact sites, and quality control.EMBO Rep. 2019 Jan;20(1):e46864. doi: 10.15252/embr.201846864. Epub 2018 Dec 10. EMBO Rep. 2019. PMID: 30530632 Free PMC article. Review.
-
Arf1 coordinates fatty acid metabolism and mitochondrial homeostasis.Nat Cell Biol. 2023 Aug;25(8):1157-1172. doi: 10.1038/s41556-023-01180-2. Epub 2023 Jul 3. Nat Cell Biol. 2023. PMID: 37400497 Free PMC article.
References
-
- Mano S, Nishimura M. Plant peroxisomes. Vitam Horm. 2005;72:111–154. full_text. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
