

Trypanosoma cruzi infection disturbs mitochondrial membrane potential and ROS production rate in cardiomyocytes
- PMID: 19686837
- PMCID: PMC2767388
- DOI: 10.1016/j.freeradbiomed.2009.08.008
Trypanosoma cruzi infection disturbs mitochondrial membrane potential and ROS production rate in cardiomyocytes
Abstract
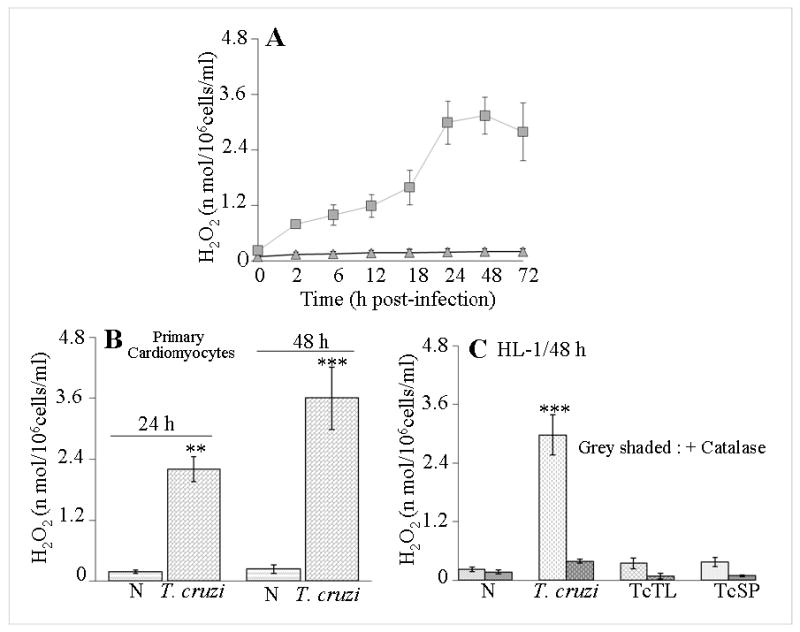
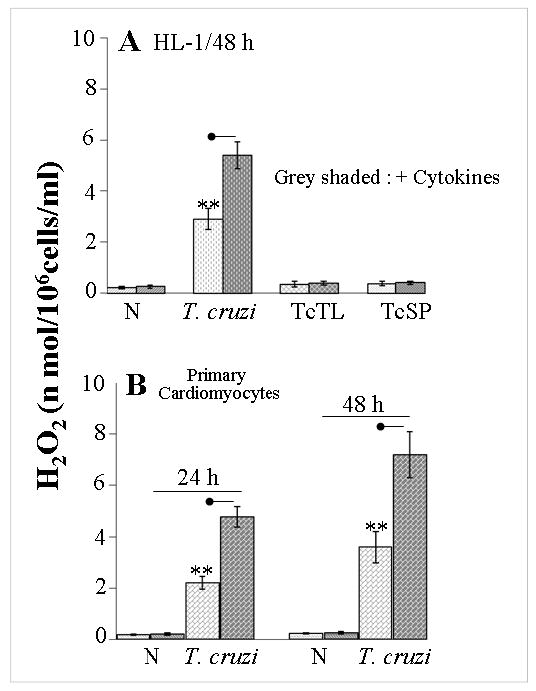
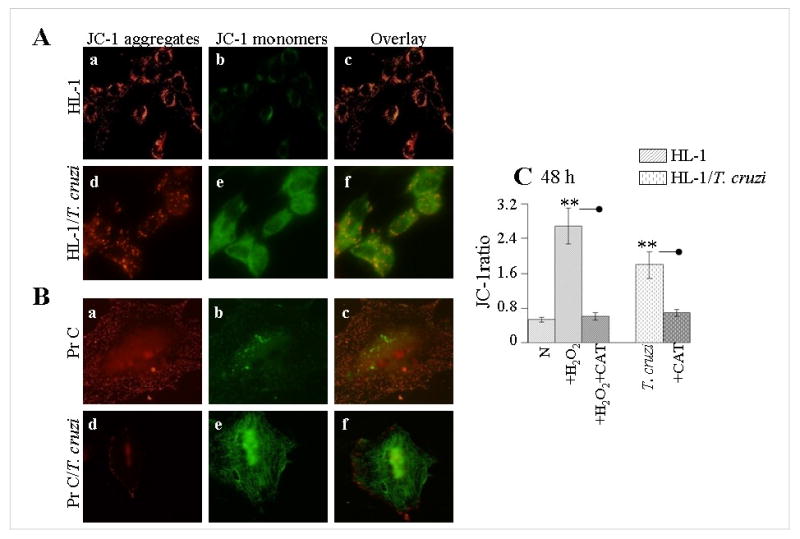
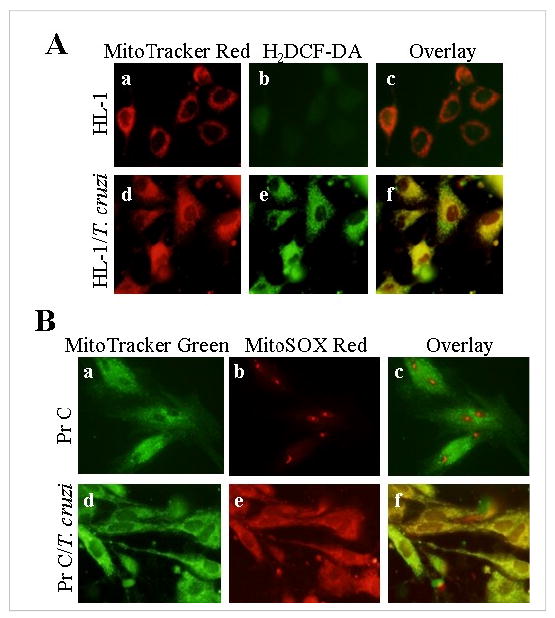
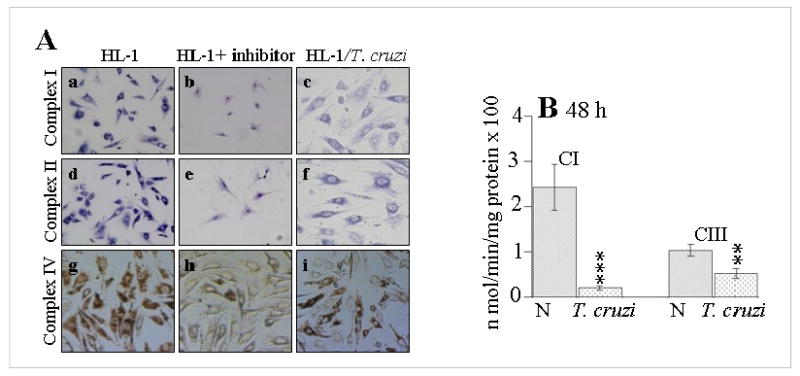
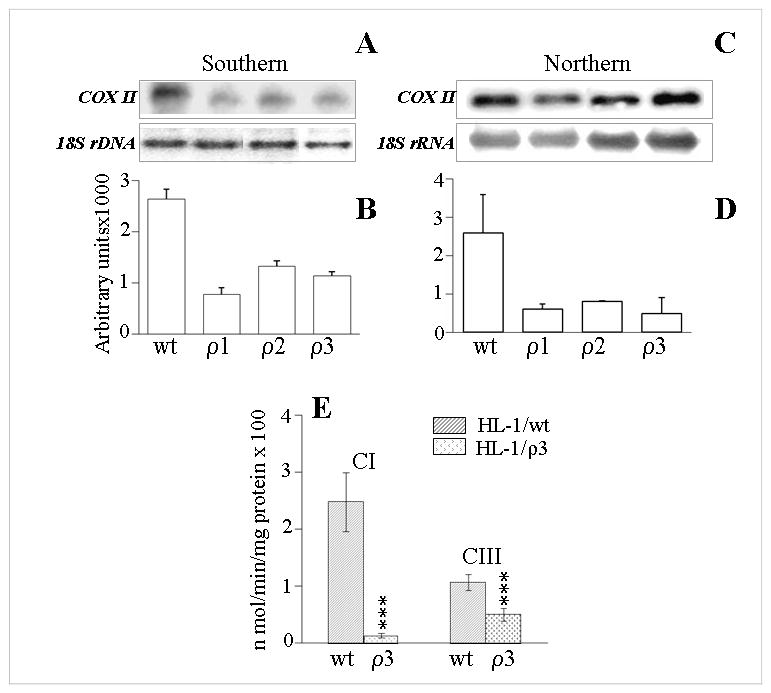
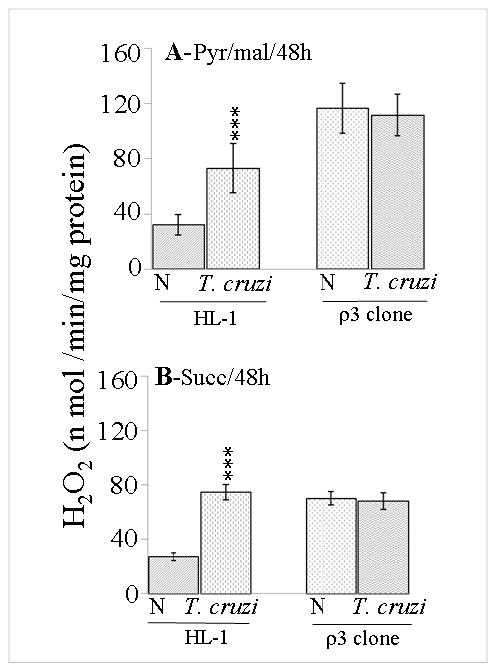
In this study, we investigated the role of Trypanosoma cruzi invasion and inflammatory processes in reactive oxygen species (ROS) production in a mouse atrial cardiomyocyte line (HL-1) and primary adult rat ventricular cardiomyocytes. Cardiomyocytes were incubated with T. cruzi (Tc) trypomastigotes, Tc lysate (TcTL), or Tc secreted proteins (TcSP) for 0-72 h, and ROS were measured by amplex red assay. Cardiomyocytes infected by T. cruzi (but not those incubated with TcTL or TcSP) exhibited a linear increase in ROS production for 2-48 h postinfection (max 18-fold increase), which was further enhanced by recombinant cytokines (IL-1beta, TNF-alpha, and IFN-gamma). We observed no increase in NADPH oxidase, xanthine oxidase, or myeloperoxidase activity, and specific inhibitors of these enzymes did not block the increased rate of ROS production in infected cardiomyocytes. Instead, the mitochondrial membrane potential was perturbed and resulted in inefficient electron transport chain (ETC) activity and enhanced electron leakage and ROS formation in infected cardiomyocytes. HL-1 rho (rho) cardiomyocytes lacked a functional ETC and exhibited no increase in ROS formation in response to T. cruzi. Together, these results demonstrate that invasion by T. cruzi and an inflammatory milieu affect mitochondrial integrity and contribute to electron transport chain inefficiency and ROS production in cardiomyocytes.
Figures
References
-
- World Health Organization. Report of the Scientific Working Group on Chagas Disease. UNDP/World Bank/WHO. 2006
-
- Higuchi MD. Endomyocardial biopsy in Chagas' heart disease: pathogenetic contributions. Sao Paulo Med J. 1995;113:821–825. - PubMed
-
- Zacks MA, Wen JJ, Vyatkina G, Bhatia V, Garg N. An overview of chagasic cardiomyopathy: pathogenic importance of oxidative stress. An Acad Bras Cienc. 2005;77:695–715. - PubMed
-
- Munoz-Fernandez MA, Fernandez MA, Fresno M. Activation of human macrophages for the killing of intracellular Trypanosoma cruzi by TNF-alpha and IFN-gamma through a nitric oxide-dependent mechanism. Immunol Lett. 1992;33:35–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
