

Different pathways for activation and deactivation in CaV1.2: a minimal gating model
- PMID: 19687230
- PMCID: PMC2737230
- DOI: 10.1085/jgp.200910272
Different pathways for activation and deactivation in CaV1.2: a minimal gating model
Abstract
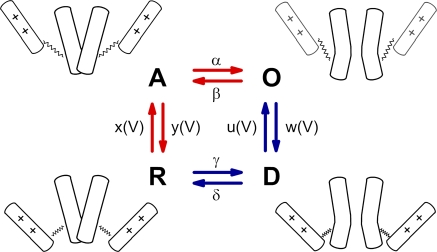
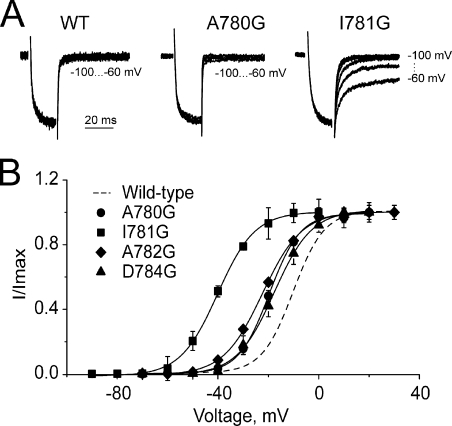
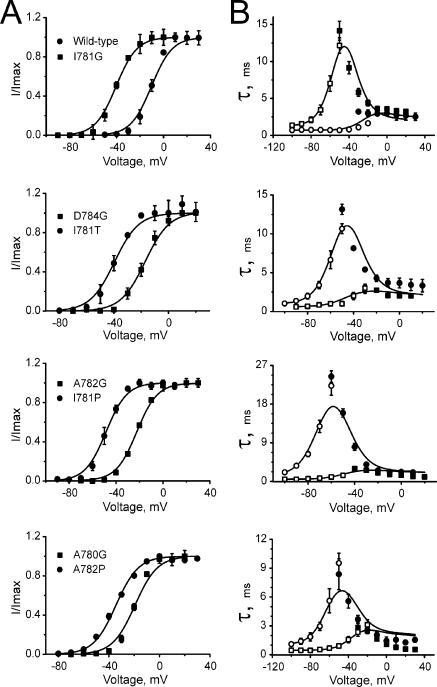
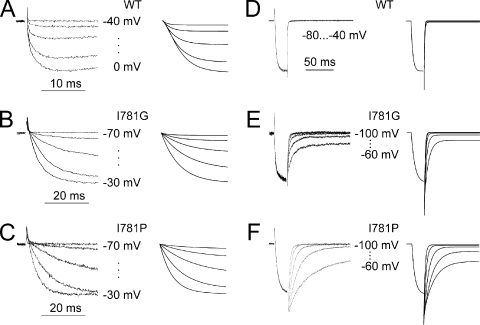
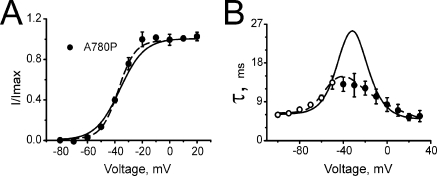
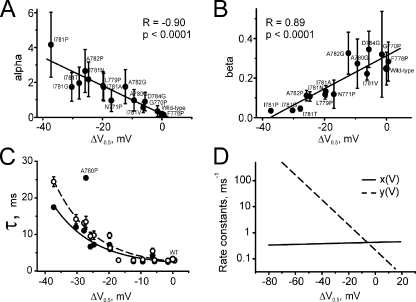
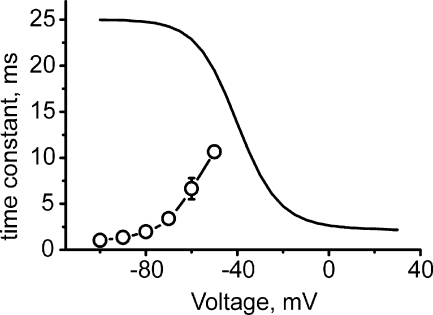
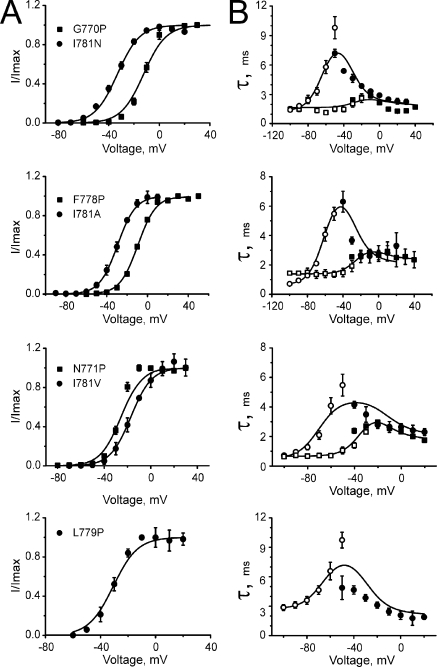
Point mutations in pore-lining S6 segments of CaV1.2 shift the voltage dependence of activation into the hyperpolarizing direction and significantly decelerate current activation and deactivation. Here, we analyze theses changes in channel gating in terms of a circular four-state model accounting for an activation R-A-O and a deactivation O-D-R pathway. Transitions between resting-closed (R) and activated-closed (A) states (rate constants x(V) and y(V)) and open (O) and deactivated-open (D) states (u(V) and w(V)) describe voltage-dependent sensor movements. Voltage-independent pore openings and closures during activation (A-O) and deactivation (D-R) are described by rate constants alpha and beta, and gamma and delta, respectively. Rate constants were determined for 16-channel constructs assuming that pore mutations in IIS6 do not affect the activating transition of the voltage-sensing machinery (x(V) and y(V)). Estimated model parameters of 15 CaV1.2 constructs well describe the activation and deactivation processes. Voltage dependence of the "pore-releasing" sensor movement ((x(V)) was much weaker than the voltage dependence of "pore-locking" sensor movement (y(V)). Our data suggest that changes in membrane voltage are more efficient in closing than in opening CaV1.2. The model failed to reproduce current kinetics of mutation A780P that was, however, accurately fitted with individually adjusted x(V) and y(V). We speculate that structural changes induced by a proline substitution in this position may disturb the voltage-sensing domain.
Figures
References
-
- Barlow D.J., Thornton J.M. 1988. Helix geometry in proteins.J. Mol. Biol. 201:601–619 - PubMed
-
- Bates D.M., Watts D.G. 1988. Nonlinear Regression Analysis and Its Application. Wiley Series in Probability and Mathematical Statistics, Inc. John Wiley and Sons, New York: 384 pp
-
- Catterall W.A. 2000. Structure and regulation of voltage-gated Ca2+ channels.Annu. Rev. Cell Dev. Biol. 16:521–555 - PubMed
-
- Cordes F.S., Bright J.N., Sansom M.S. 2002. Proline-induced distortions of transmembrane helices.J. Mol. Biol. 323:951–960 - PubMed
-
- Doyle D.A., Morais Cabral J., Pfuetzner R.A., Kuo A., Gulbis J.M., Cohen S.L., Chait B.T., MacKinnon R. 1998. The structure of the potassium channel: molecular basis of K+ conduction and selectivity.Science. 280:69–77 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
