

Activated Abl kinase inhibits oncogenic transforming growth factor-beta signaling and tumorigenesis in mammary tumors
- PMID: 19690215
- PMCID: PMC2812049
- DOI: 10.1096/fj.09-138412
Activated Abl kinase inhibits oncogenic transforming growth factor-beta signaling and tumorigenesis in mammary tumors
Abstract
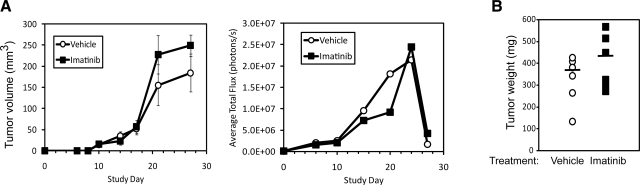
Transforming growth factor-beta (TGF-beta) is a ubiquitous cytokine with dual roles in tumor suppression and promotion, and these dichotomous functions have frustrated the development of therapies targeting oncogenic signaling by TGF-beta. In comparison, Abl is well established as an initiator of hematopoietic cancers; however, a clear role for Abl in regulating solid tumor development remains elusive. Here, we investigated the role of Abl in TGF-beta-mediated epithelial-mesenchymal transition (EMT) in normal and metastatic mammary epithelial cells (MECs). In doing so, we identified Abl as an essential regulator of MEC morphology and showed that Abl inactivation was sufficient to induce phenotypic and transcriptional EMT in normal MECs. Increasing Abl activity in metastatic MECs resulted in their complete morphological reversion, restored their cytostatic response to TGF-beta, and blocked their secretion of matrix metalloproteinases induced by TGF-beta. Constitutively active Abl expression blocked TGF-beta-responsive mammary tumor growth in mice, while Imatinib therapy afforded no clinical benefit in mice bearing mammary tumors. Collectively, this investigation establishes Abl as a potent mediator of MEC identity, and as a suppressor of oncogenic TGF-beta signaling during mammary tumorigenesis. Notably, our findings strongly caution against the use of pharmacological Abl antagonists in the treatment of developing and progressing mammary tumors.
Figures
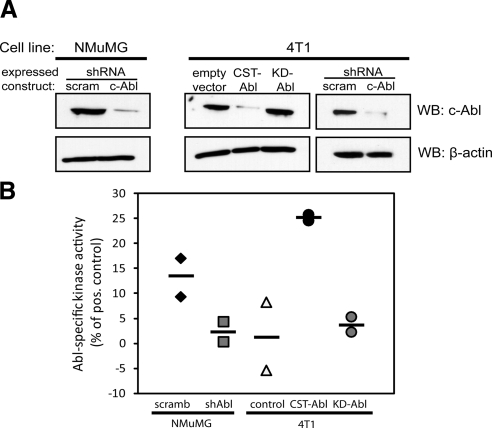
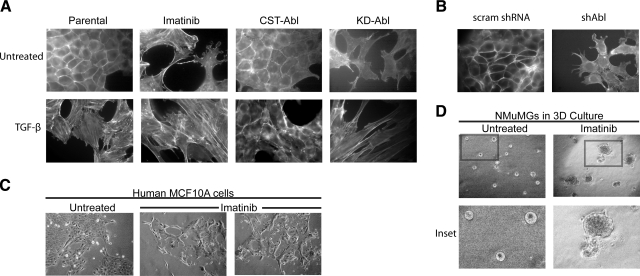
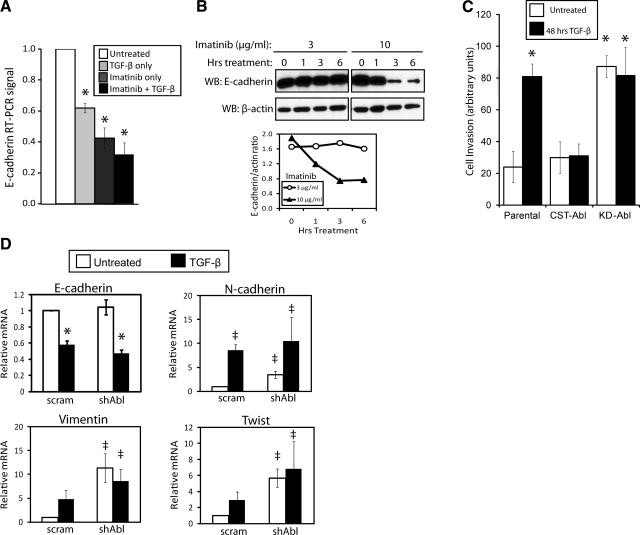
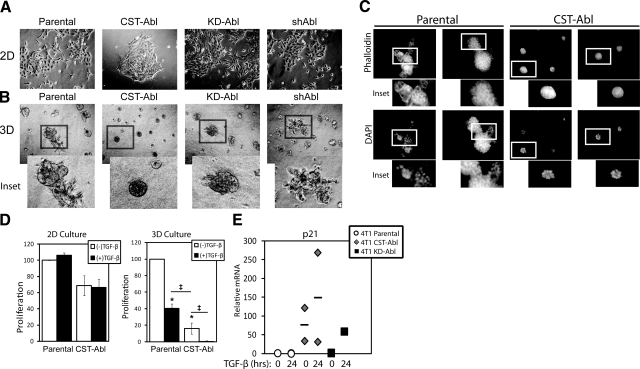
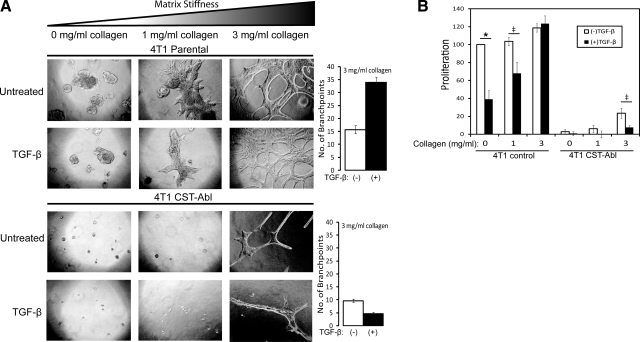
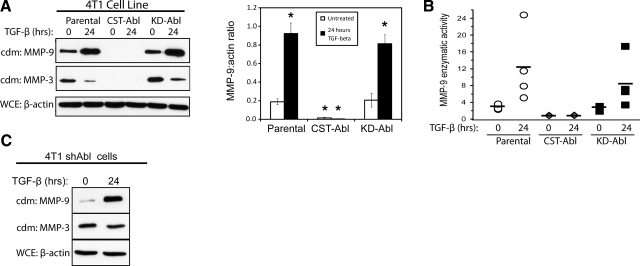
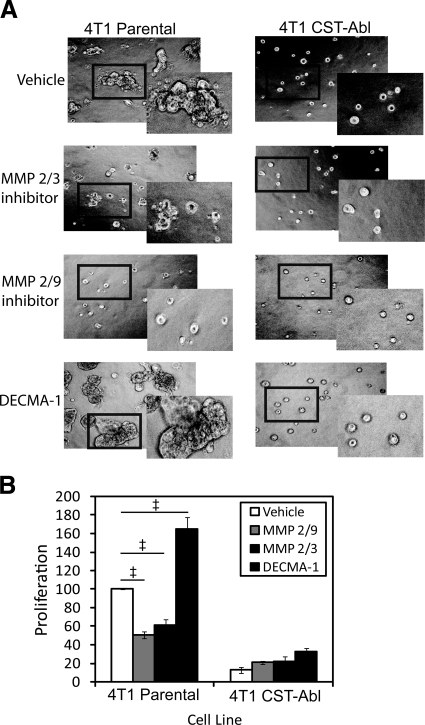
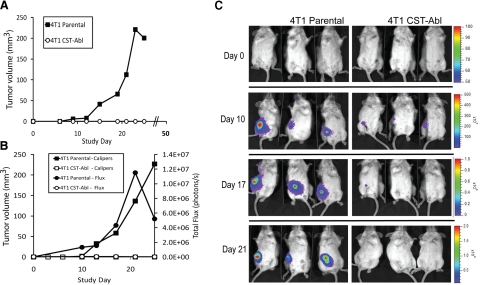
References
-
- Benson J R. Role of TGF-β in breast carcinogenesis. Lancet Oncol. 2004;5:229–239. - PubMed
-
- Pardali K, Moustakas A. Actions of TGF-β as tumor suppressor and pro-metastatic factor in human cancer. Biochim Biophys Acta. 2007;1775:21–62. - PubMed
-
- Wakefield L M, Piek E, Bottinger E P. TGF-β signaling in mammary gland development and tumorigenesis. J Mammary Gland Biol Neoplasia. 2001;6:67–82. - PubMed
-
- Wakefield L M, Roberts A B. TGF-β signaling: positive and negative effects on tumorigenesis. Curr Opin Genet Dev. 2002;12:22–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
