

Structure-function studies of claudin extracellular domains by cysteine-scanning mutagenesis
- PMID: 19690347
- PMCID: PMC2781464
- DOI: 10.1074/jbc.M109.043752
Structure-function studies of claudin extracellular domains by cysteine-scanning mutagenesis
Abstract
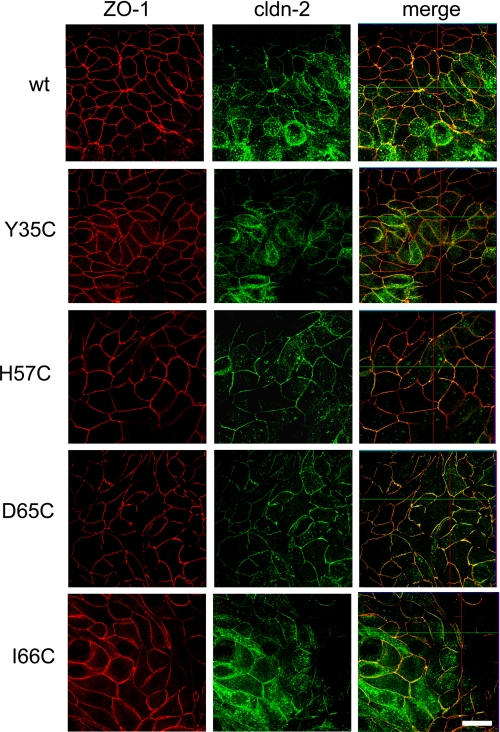
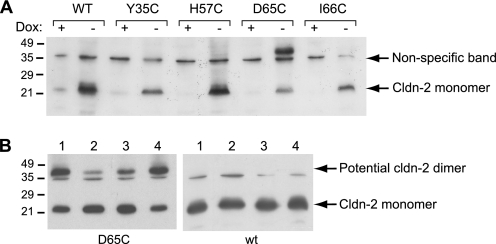
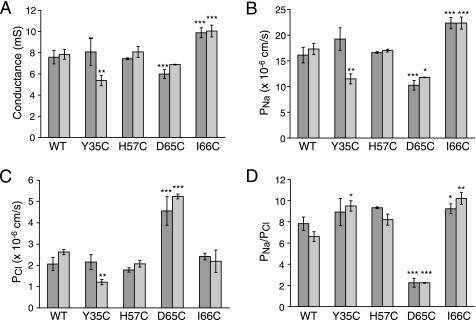
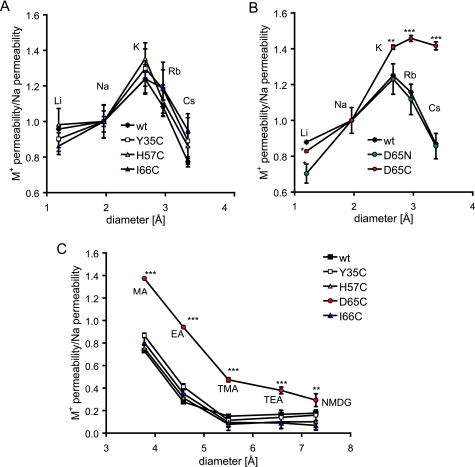
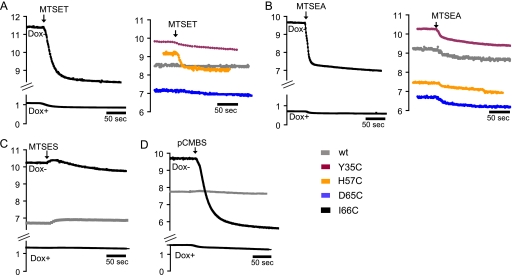
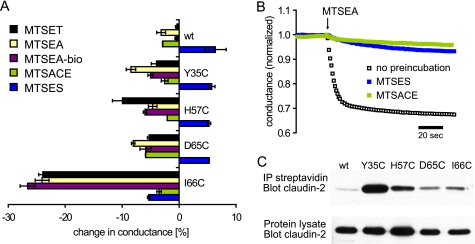
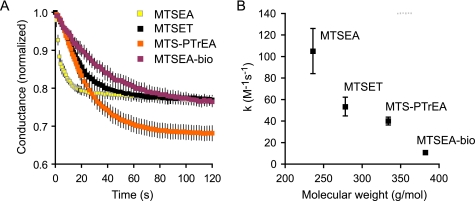
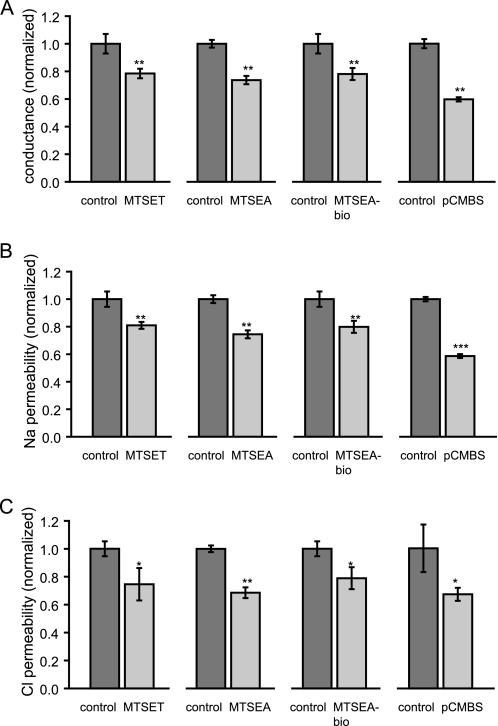
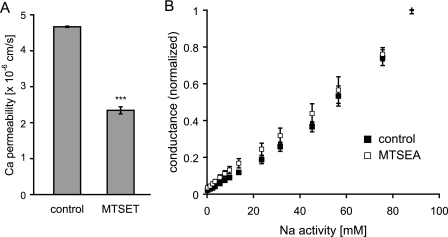
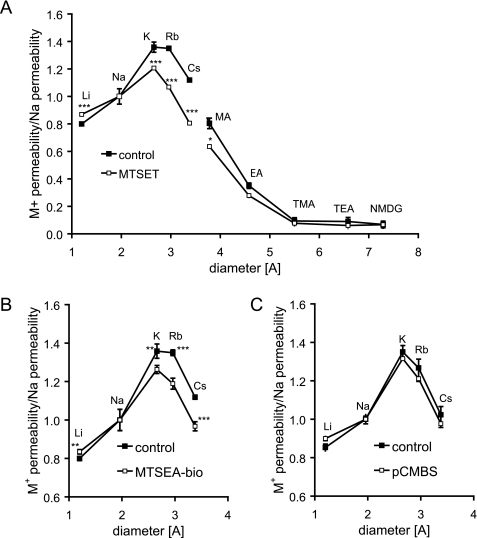
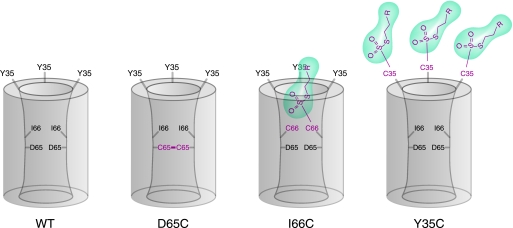
Claudins form size- and charge-selective pores in the tight junction that control the paracellular flux of inorganic ions and small molecules. However, the structural basis for ion selectivity of paracellular pores is poorly understood. Here we applied cysteine scanning to map the paracellular pathway of ion permeation across claudin-2-transfected Madin-Darby canine kidney type I cells. Four potential pore-lining amino acid residues in the first extracellular loop were mutated to cysteine and screened for their accessibility to thiol-reactive reagents. All mutants were functional except D65C, which formed dimers by intermolecular disulfide bonding, leading to a loss of charge and size selectivity. This suggests that claudin-2 pores are multimeric and that Asp(65) lies close to a protein-protein interface. Methanethiosulfonate reagents of different size and charge and the organic mercury derivate, p-(chloromercuri)benzenesulfonic acid, significantly decreased paracellular ion permeation across I66C-transfected cells by a mechanism that suggests steric blocking of the pore. The conductance of wild-type claudin-2 and the other cysteine mutants was only weakly affected. The rate of reaction with I66C decreased dramatically with increasing size of the reagent, suggesting that Ile(66) is buried deep within a narrow segment of the pore with its side group facing into the lumen. Furthermore, labeling with N-biotinoylaminoethyl methanethiosulfonate showed that I66C was weakly reactive, whereas Y35C was strongly reactive, suggesting that Tyr(35) is located at the protein surface outside of the pore.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
