

Aldosterone mediates activation of the thiazide-sensitive Na-Cl cotransporter through an SGK1 and WNK4 signaling pathway
- PMID: 19690383
- PMCID: PMC2735908
- DOI: 10.1172/JCI38323
Aldosterone mediates activation of the thiazide-sensitive Na-Cl cotransporter through an SGK1 and WNK4 signaling pathway
Abstract
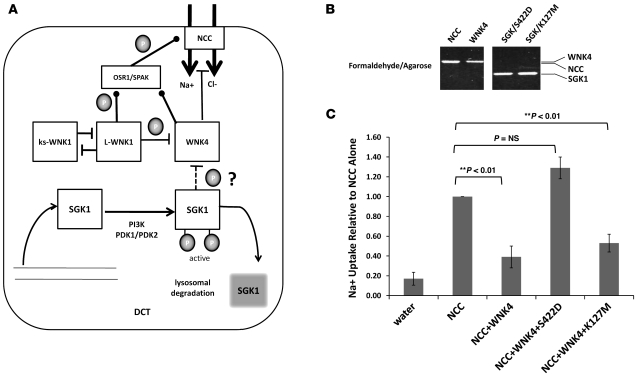
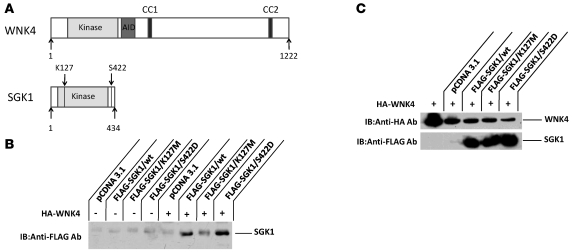
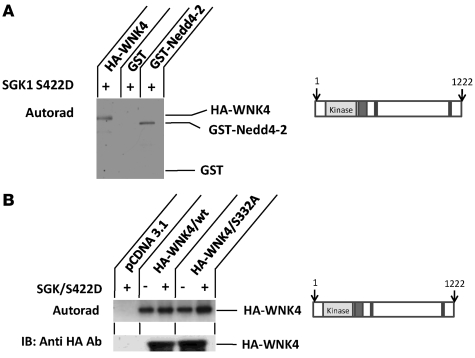
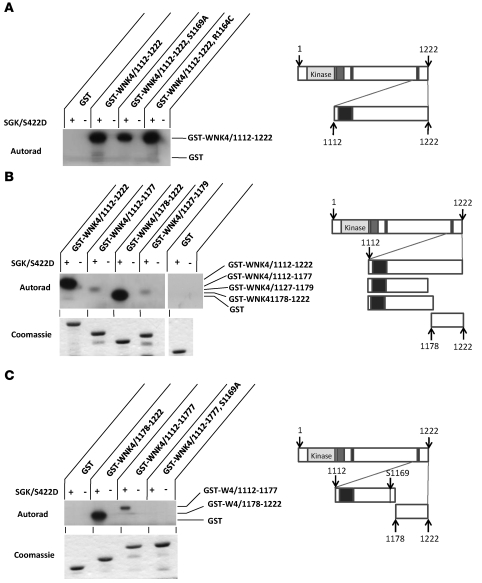
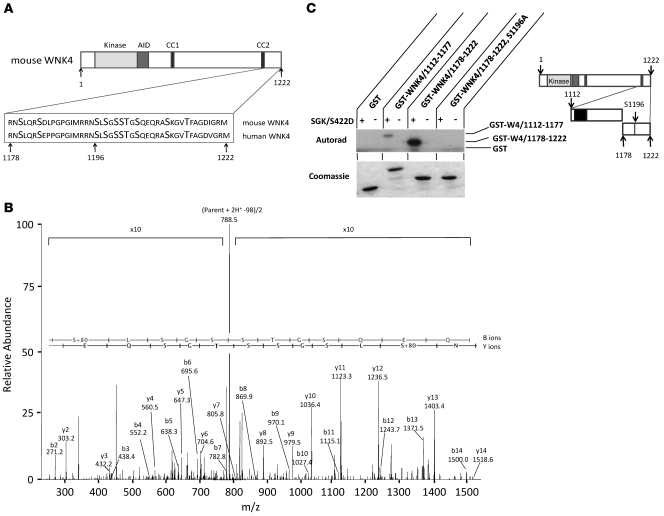
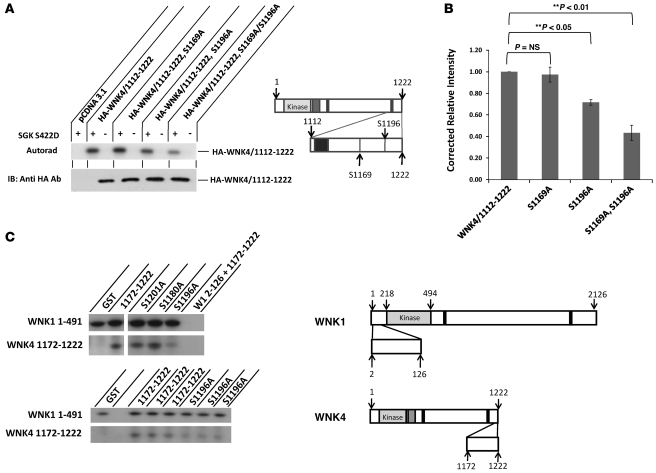
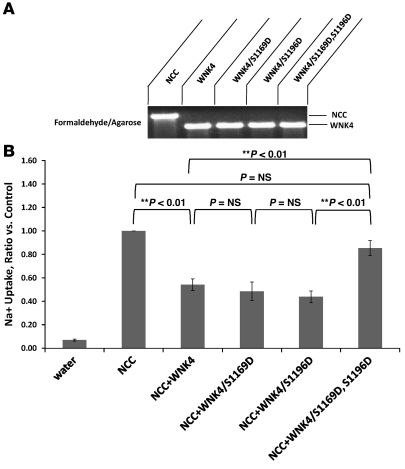
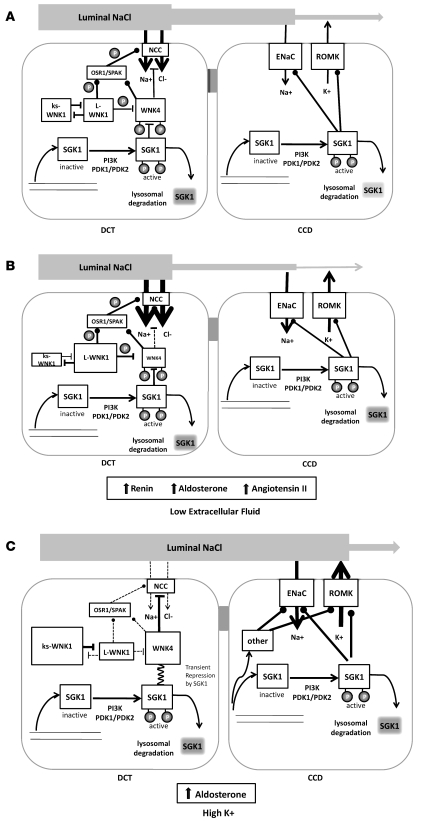
Aldosterone regulates volume homeostasis and blood pressure by enhancing sodium reabsorption in the kidney's distal nephron (DN). On the apical surface of these renal epithelia, aldosterone increases expression and activity of the thiazide-sensitive Na-Cl cotransporter (NCC) and the epithelial sodium channel (ENaC). While the cellular mechanisms by which aldosterone regulates ENaC have been well characterized, the molecular mechanisms that link aldosterone to NCC-mediated Na+/Cl- reabsorption remain elusive. The serine/threonine kinase with-no-lysine 4 (WNK4) has previously been shown to reduce cell surface expression of NCC. Here we measured sodium uptake in a Xenopus oocyte expression system and found that serum and glucocorticoid-induced kinase 1 (SGK1), an aldosterone-responsive gene expressed in the DN, attenuated the inhibitory effect of WNK4 on NCC activity. In addition, we showed--both in vitro and in a human kidney cell line--that SGK1 bound and phosphorylated WNK4. We found one serine located within an established SGK1 consensus target sequence, and the other within a motif that was, to our knowledge, previously uncharacterized. Mutation of these target serines to aspartate, in order to mimic phosphorylation, attenuated the effect of WNK4 on NCC activity in the Xenopus oocyte system. These data thus delineate what we believe to be a novel mechanism for aldosterone activation of NCC through SGK1 signaling of WNK4 kinase.
Figures
References
-
- Reilly R.F., Ellison D.H. Mammalian distal tubule: physiology, pathophysiology, and molecular anatomy. Physiol. Rev. 2000;80:277–313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
