

Dbf4-Cdc7 phosphorylation of Mcm2 is required for cell growth
- PMID: 19692334
- PMCID: PMC2781428
- DOI: 10.1074/jbc.M109.039123
Dbf4-Cdc7 phosphorylation of Mcm2 is required for cell growth
Abstract
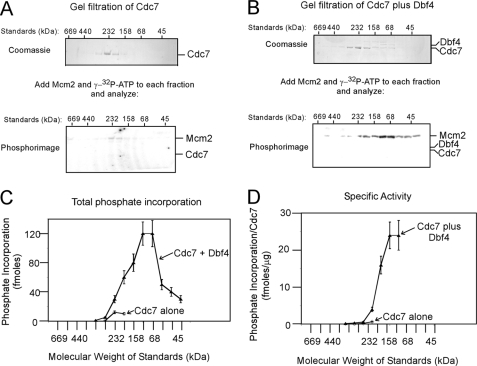
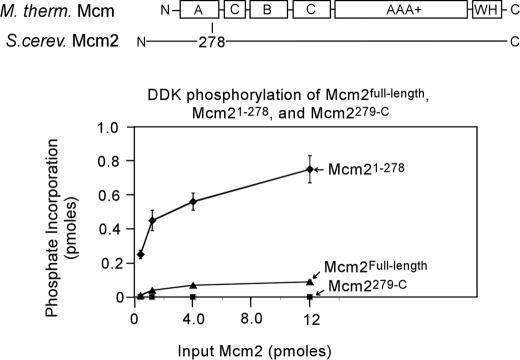
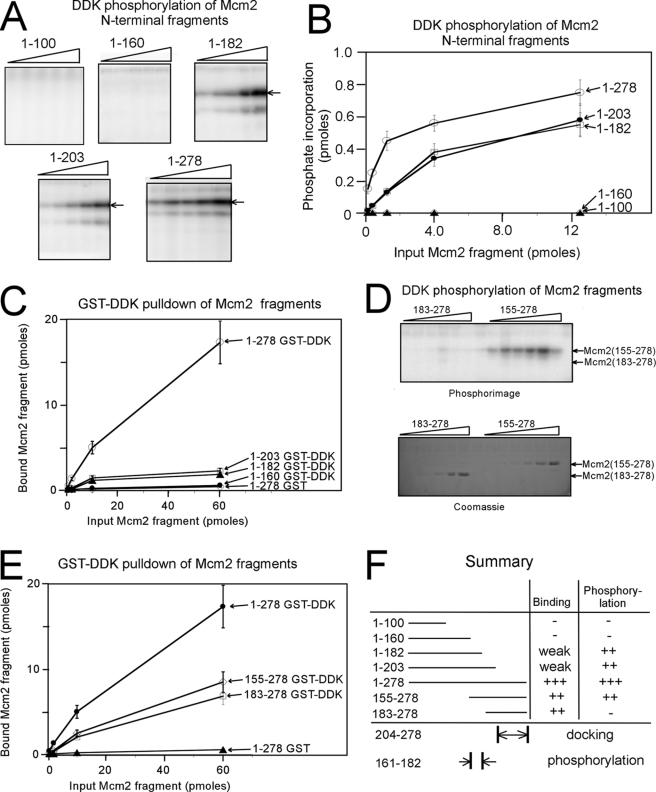
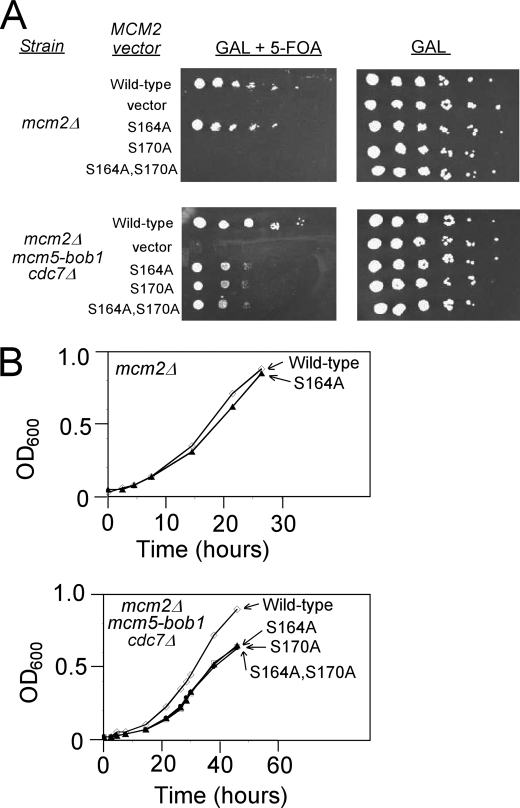
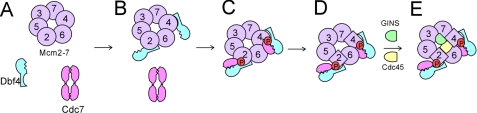
The Dbf4-Cdc7 kinase (DDK) is required for the activation of the origins of replication, and DDK phosphorylates Mcm2 in vitro. We find that budding yeast Cdc7 alone exists in solution as a weakly active multimer. Dbf4 forms a likely heterodimer with Cdc7, and this species phosphorylates Mcm2 with substantially higher specific activity. Dbf4 alone binds tightly to Mcm2, whereas Cdc7 alone binds weakly to Mcm2, suggesting that Dbf4 recruits Cdc7 to phosphorylate Mcm2. DDK phosphorylates two serine residues of Mcm2 near the N terminus of the protein, Ser-164 and Ser-170. Expression of mcm2-S170A is lethal to yeast cells that lack endogenous MCM2 (mcm2Delta); however, this lethality is rescued in cells harboring the DDK bypass mutant mcm5-bob1. We conclude that DDK phosphorylation of Mcm2 is required for cell growth.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
