

An N-terminal addressing sequence targets NLRX1 to the mitochondrial matrix
- PMID: 19692591
- PMCID: PMC2871076
- DOI: 10.1242/jcs.051193
An N-terminal addressing sequence targets NLRX1 to the mitochondrial matrix
Abstract
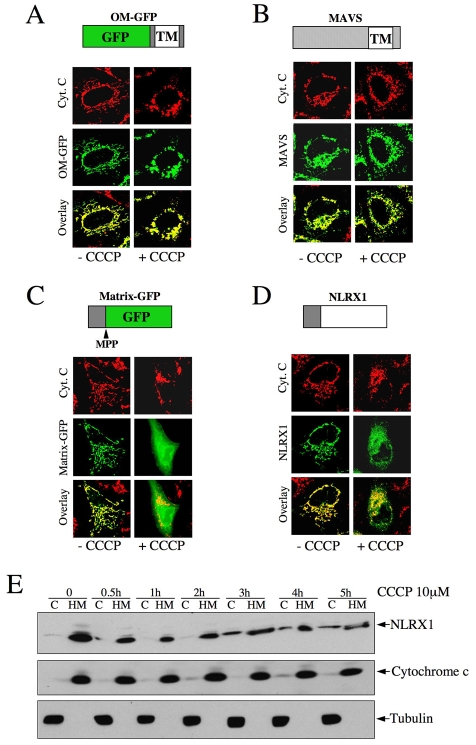
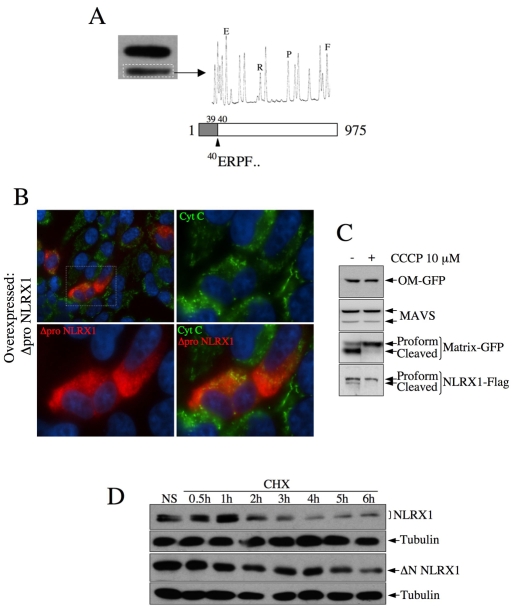
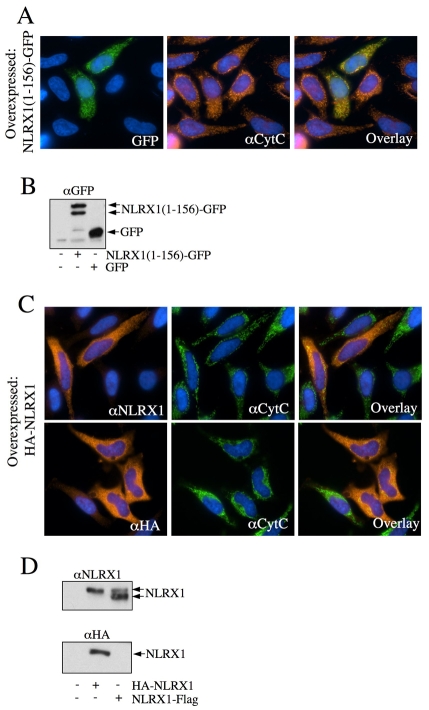
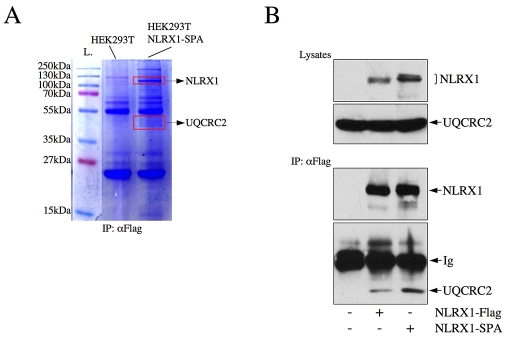
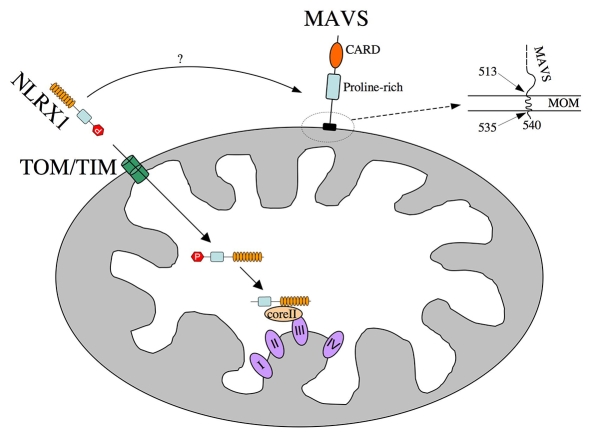
NLRX1 is the only member of the Nod-like receptor (NLR) family that is targeted to the mitochondria, and its overexpression induces the generation of reactive oxygen species (ROS), thus impacting on NFkappaB- and JNK-dependent signaling cascades. In addition, NLRX1 has been shown to interact with MAVS (also known as IPS-1, VISA and Cardif) at the mitochondrial outer membrane and to modulate antiviral responses. Here we report that NLRX1 has a functional leader sequence and fully translocates to the mitochondrial matrix via a mechanism requiring the mitochondrial inner-membrane potential, DeltaPsim. Importantly, we failed to detect NLRX1 at the mitochondrial outer membrane. We also show that the leader sequence of NLRX1 is removed, which generates a mature protein lacking the first 39 amino acids through a maturation process that is common for mitochondrial-matrix proteins. Finally, we identified UQCRC2, a matrix-facing protein of the respiratory chain complex III, as an NLRX1-interacting molecule, thus providing a molecular basis for the role of NLRX1 in ROS generation. These results provide the first identification of a protein belonging to the NLR family that is targeted to the mitochondrial matrix.
Figures
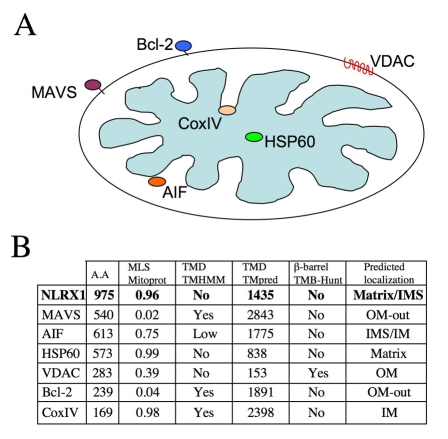
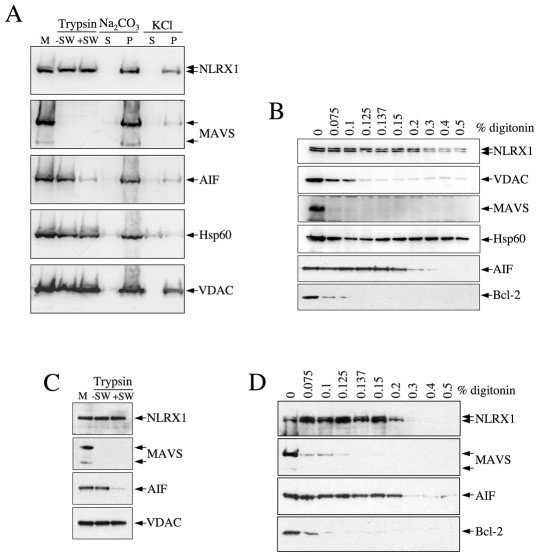
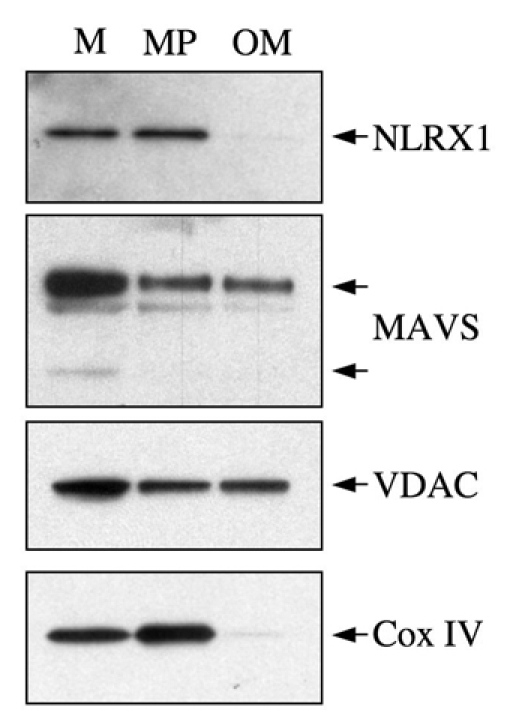
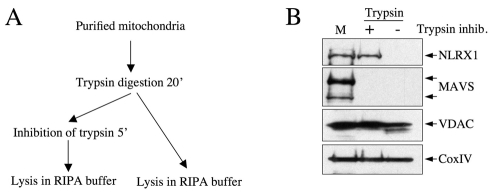
References
-
- Akira, S., Uematsu, S. and Takeuchi, O. (2006). Pathogen recognition and innate immunity. Cell 124, 783-801. - PubMed
-
- Arnoult, D. (2008). Apoptosis-associated mitochondrial outer membrane permeabilization assays. Methods 44, 229-234. - PubMed
-
- Bell, E. L., Klimova, T. A., Eisenbart, J., Moraes, C. T., Murphy, M. P., Budinger, G. R. and Chandel, N. S. (2007). The Qo site of the mitochondrial complex III is required for the transduction of hypoxic signaling via reactive oxygen species production. J. Cell Biol. 177, 1029-1036. - PMC - PubMed
-
- Benko, S., Philpott, D. J. and Girardin, S. E. (2008). The microbial and danger signals that activate Nod-like receptors. Cytokine 43, 368-373. - PubMed
-
- Fritz, J. H., Ferrero, R. L., Philpott, D. J. and Girardin, S. E. (2006). Nod-like proteins in immunity, inflammation and disease. Nat. Immunol. 7, 1250-1257. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
