

Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment
- PMID: 19693011
- PMCID: PMC2931797
- DOI: 10.1038/nature08268
Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment
Abstract
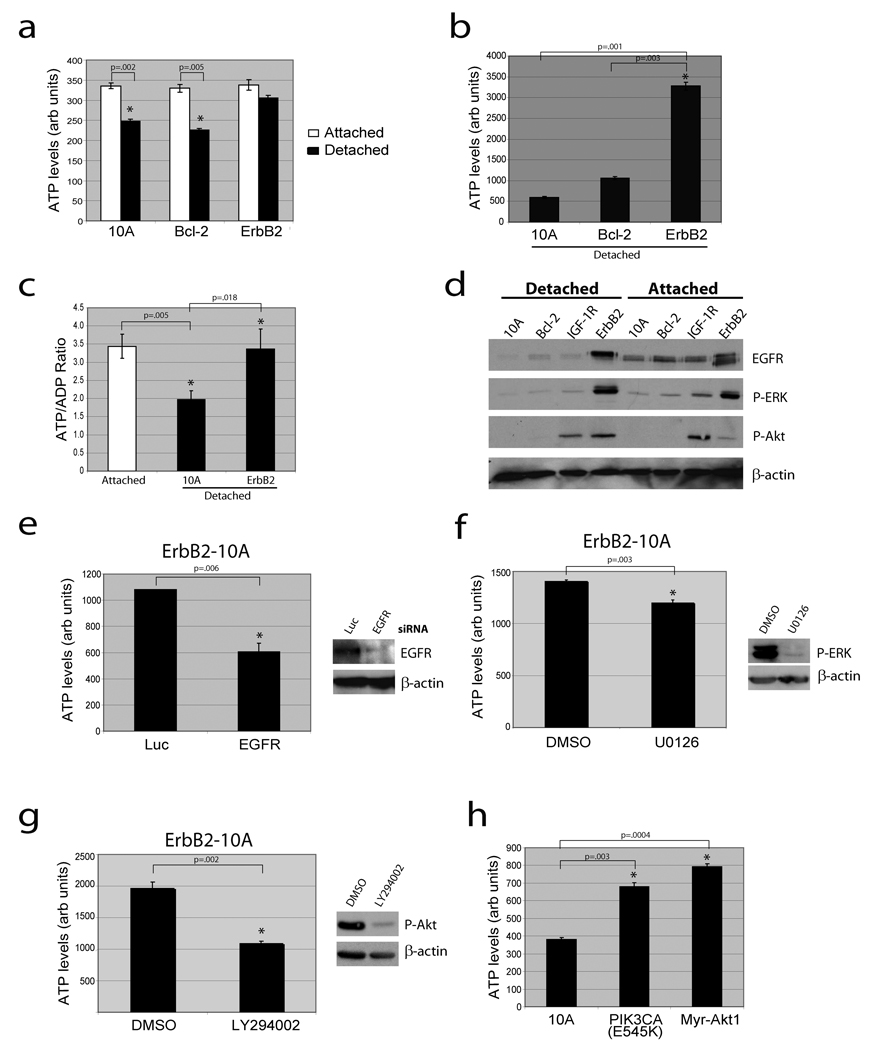
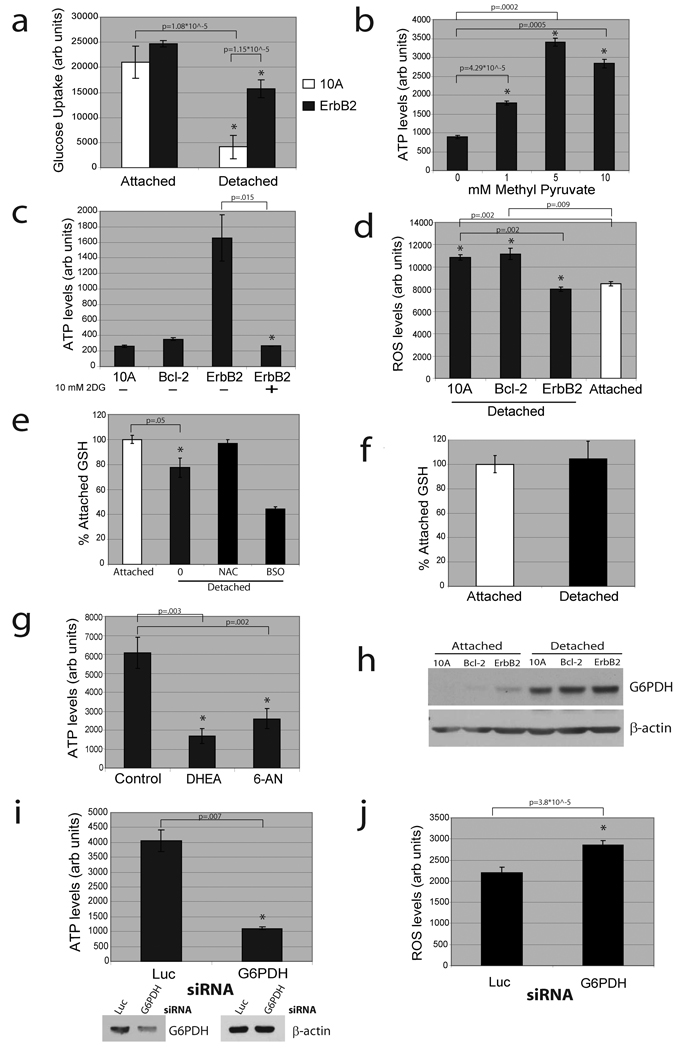
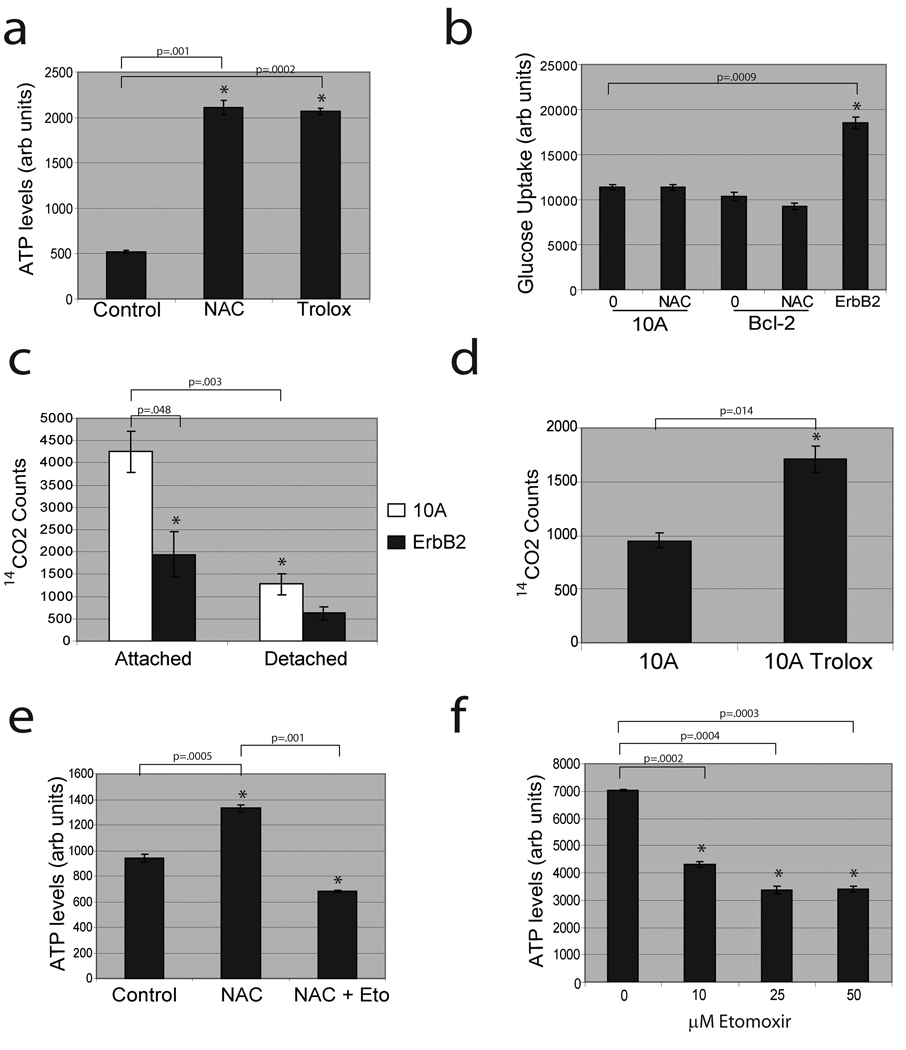
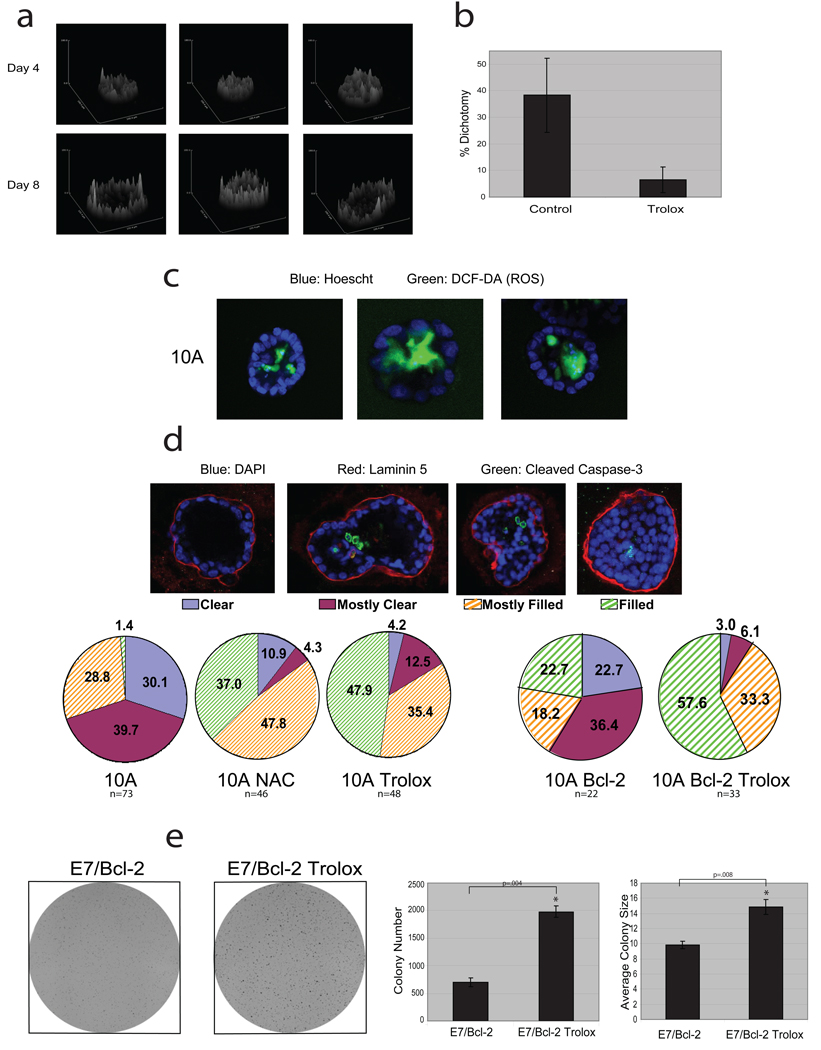
Normal epithelial cells require matrix attachment for survival, and the ability of tumour cells to survive outside their natural extracellular matrix (ECM) niches is dependent on acquisition of anchorage independence. Although apoptosis is the most rapid mechanism for eliminating cells lacking appropriate ECM attachment, recent reports suggest that non-apoptotic death processes prevent survival when apoptosis is inhibited in matrix-deprived cells. Here we demonstrate that detachment of mammary epithelial cells from ECM causes an ATP deficiency owing to the loss of glucose transport. Overexpression of ERBB2 rescues the ATP deficiency by restoring glucose uptake through stabilization of EGFR and phosphatidylinositol-3-OH kinase (PI(3)K) activation, and this rescue is dependent on glucose-stimulated flux through the antioxidant-generating pentose phosphate pathway. Notably, we found that the ATP deficiency could be rescued by antioxidant treatment without rescue of glucose uptake. This rescue was found to be dependent on stimulation of fatty acid oxidation, which is inhibited by detachment-induced reactive oxygen species (ROS). The significance of these findings was supported by evidence of an increase in ROS in matrix-deprived cells in the luminal space of mammary acini, and the discovery that antioxidants facilitate the survival of these cells and enhance anchorage-independent colony formation. These results show both the importance of matrix attachment in regulating metabolic activity and an unanticipated mechanism for cell survival in altered matrix environments by antioxidant restoration of ATP generation.
Figures
Comment in
-
Cancer: The fat and the furious.Nature. 2009 Sep 3;461(7260):44-5. doi: 10.1038/461044a. Nature. 2009. PMID: 19727186 No abstract available.
References
-
- Simpson CD, Anyiwe K, Schimmer AD. Anoikis resistance and tumor metastasis. Cancer Lett. 2008 - PubMed
-
- Frisch SM, Screaton RA. Anoikis mechanisms. Curr Opin Cell Biol. 2001;13:555–562. - PubMed
-
- Debnath J, et al. The role of apoptosis in creating and maintaining luminal space within normal and oncogene-expressing mammary acini. Cell. 2002;111:29–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
