

Review
doi: 10.1038/nrm2748.
Endocytosis and signalling: intertwining molecular networks
Affiliations
- PMID: 19696798
- PMCID: PMC2895425
- DOI: 10.1038/nrm2748
Item in Clipboard
Review
Endocytosis and signalling: intertwining molecular networks
Nat Rev Mol Cell Biol.
2009 Sep.
Abstract
Cell signalling and endocytic membrane trafficking have traditionally been viewed as distinct processes. Although our present understanding is incomplete and there are still great controversies, it is now recognized that these processes are intimately and bidirectionally linked in animal cells. Indeed, many recent examples illustrate how endocytosis regulates receptor signalling (including signalling from receptor tyrosine kinases and G protein-coupled receptors) and, conversely, how signalling regulates the endocytic pathway. The mechanistic and functional principles that underlie the relationship between signalling and endocytosis in cell biology are becoming increasingly evident across many systems.
Figures
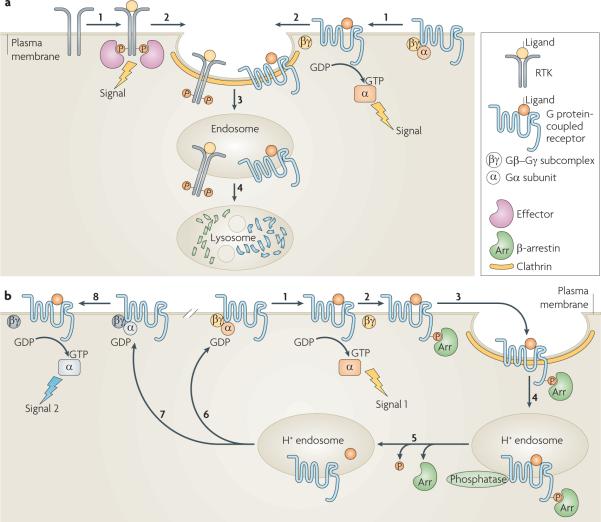
a | Schematic depicting signal attenuation. Ligand-induced activation of receptor tyrosine kinases (RTKs) or G protein-coupled receptors (GPCRs) promotes signalling from the plasma membrane by the receptor-mediated recruitment of signalling effectors to phosphorylated RTKs, or the activation of G proteins associated with GPCRs when the Gα subunit is bound to GTP (step 1). GPCR signalling can be mediated both by the GTP-bound Gα subunit (as depicted) and by the Gβ–Gγ subcomplex. Receptor recruitment into coated pits (step 2) and clathrin-dependent endocytosis (step 3) attenuate signalling by separating the receptors from plasma membrane-delimited substrates and/or mediators. Some receptors traffic to lysosomes after endocytosis, which results in their downregulation by proteolysis and further attenuates signalling (step 4). b | Schematic depicting signal desensitization, re-sensitization and pathway switching. The ability of some GPCRs to activate trimeric G proteins (signal 1; step 1) is attenuated before endocytosis by receptor phosphorylation and β-arrestin binding (step 2). Such desensitized receptor–β-arrestin complexes concentrate into clathrin-coated pits (step 3) and are endocytosed into acidic (H+) endosomes (step 4), which promotes various events that may include (depending on the receptor and cell type) ligand dissociation or destruction, dissociation of β-arrestin and phosphatase-catalysed dephosphorylation (step 5). Recycling (step 6) restores the receptors to the cell surface, re-sensitizing the cell for another round of signalling. In some cases GPCR recycling inserts receptors into a different G protein-containing environment (step 7), which produces a `switch' in signalling specificity following subsequent receptor activation (signal 2; step 8).
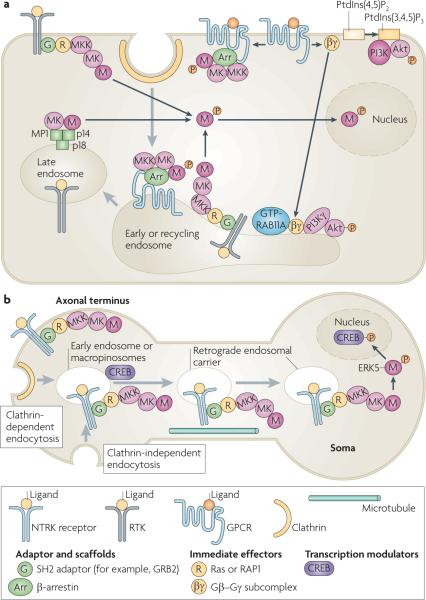
a | Activation of mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)–Akt signalling cascades by receptor tyrosine kinases (RTKs) and G protein-coupled receptors (GPCRs) occurs at the plasma membrane and in early, recycling and late endosomes. In all of these locations activated, phosphorylated MAPK dissociates from the MAPK kinase (MK), unless stably associated with a scaffold protein such as β-arrestin, and phosphorylates substrates in the cytoplasm and nucleus. When GPCRs are stimulated by lysophosphatidic acid, the activated G protein subcomplex Gβ–Gγ can activate PI3K and Akt at the plasma membrane and also translocate to the early and recycling endosomes to activate endosomal PI3K γ-isoform, which leads to the phosphorylation and activation of Akt in endosomes. b | Model of retrograde endosome signalling in neurons. Ligand-bound nerve growth factor receptors (NTRKs) are internalized by clathrin-dependent and clathrin-independent endocytosis, with associated components of the MAPK signalling cascade, into early endosomes or macropinosomes that have a multivesicular body-like morphology. NTRK signalling complexes are delivered to the soma in retrograde endosomal carriers, which are a population of early or late endosomes and macropinosomes, by dynein motor-mediated microtubular transport. The MAPK extracellular signal-regulated kinase 5 (ERK5) is then phosphorylated and activated in the soma. It phosphorylates the cAMP responsive element-binding protein (CREB), which regulates the transcription of anti-apoptotic genes. Grey arrows show trafficking pathways. Black arrows show protein translocations or complex assembly. GRB2, growth factor receptor-bound protein 2; MP1, MEK1 partner 1; PtdIns(3,4,5)P3, phosphatidylinositol-3,4,5-trisphosphate; PtdIns(4,5)P2, phosphatidylinositol-4,5-bisphosphate; SH2, Src homology 2.
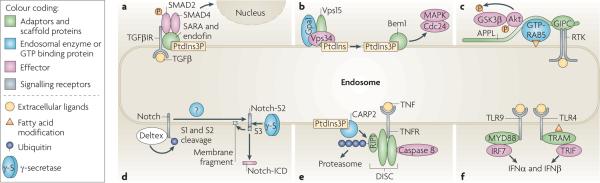
a | Endosomal recruitment of SMAD2 and SMAD4 by the FYVE domain adaptors SARA (SMAD anchor for receptor activation) and endofin, respectively, allows efficient phosphorylation of SMAD2 by internalized transforming growth factor-β receptor (TGFβR) and the formation of active SMAD2–SMAD4 complexes. b | Following the activation of the yeast protein Ste2, the GTP-binding α1 subunit (Gpa1) of the trimeric G protein complex binds to the endosomal protein Vps15. This activates the yeast phosphoinositide 3-kinase Vps34, which converts phosphatidylinositol (PtdIns) to phosphatidylinositol-3-phosphate (PtdIns3P). The FYVE domain-containing protein bud emergence protein 1 (Bem1) is subsequently recruited to PtdIns3P, where it potentiates the activation of Cdc42 and mitogen-activated protein kinase (MAPK). c | APPL1 (adaptor protein, phosphotyrosine interaction, PH domain and leucine zipper-containing 1) and APPL2 are targeted by GTP-bound RAB5 to early endosomes. They can also directly, or indirectly through GIPC (GAIP-interacting protein, C-terminus), interact with receptor tyrosine kinases (RTKs). Activated Akt and GSK3β (glycogen synthase kinase 3β) transiently associate with APPL, which leads to the phosphorylation of GSK3β in endosomes. d | Deltex promotes the ubiquitylation, endocytosis and accumulation of full-length Notch in late endosomes. The S1 and S2 Notch cleavages may be mediated by proteolytic enzymes in the late endosomes, and the Notch intracellular domain fragment (Notch-ICD) is released by γ-secretase present in late endosomes. e | Death-inducing signalling complex (DISC) is recruited to ligand-bound tumour necrosis factor receptor (TNFR). Receptor interacting protein 1 (RIP1), a component of DISC, is polyubiquitylated by the FYVE domain-containing E3 ligase CARP2 (caspases 8 and 10-associated RING finger protein) in endosomes and degraded by the proteasome, which allows the recruitment and activation of caspase 8. f | Activated Toll-like receptor 4 (TLR4) binds to the adaptor complex formed by myristoylated TRIF-related adaptor molecule (TRAM) and TIR-domain-containing adaptor protein inducing IFNβ (TRIF) only in endosomes, which allows the transcription of type I interferon (IFN) genes. Sustained interaction of ligand–TLR9 complexes with the adaptor protein myeloid differentiation primary response protein 88 (MYD88) and IFN regulatory factor 7 (IRF7) in endosomes is necessary for a potent type I IFN response.
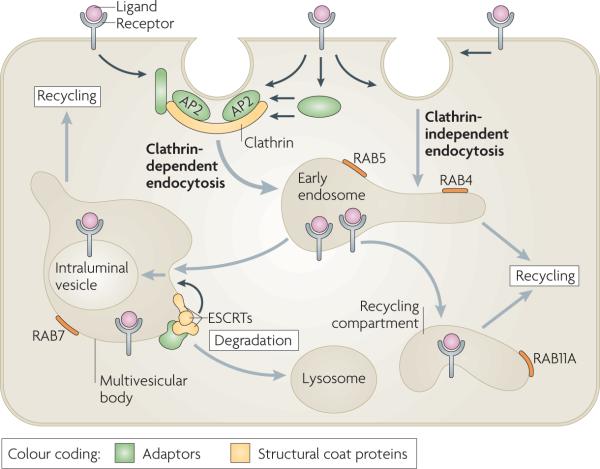
Endocytosis involves the capture of transmembrane proteins and their extracellular ligands into cytoplasmic vesicles that are pinched off from the plasma membrane (see the figure). The best-studied pathway of receptor internalization is mediated by clathrin-coated pits. These are small areas of the plasma membrane that are covered from the cytoplasmic surface with clathrin triskelions, which consist of three clathrin heavy chains and three clathrin light chains assembled into the polyhedral clathrin lattice. Receptors are recruited to clathrin-coated pits by directly interacting with the clathrin coat adaptor complex AP2 or by binding to other adaptor proteins, which in turn interacts with the clathrin heavy chain and/or AP2. Clathrin-coated pits invaginate inwards with the help of several accessory proteins and pinch off to form a clathrin-coated vesicle in a process that requires the GTPase dynamin. Several clathrin-independent pathways of endocytosis also exist, although the precise mechanisms and structural components involved in these pathways are not well understood. Endocytic vesicles derived from both clathrin-dependent and clathrin-independent endocytosis fuse with early endosomes. Endosomal trafficking is controlled by several Rab proteins — small GTP-binding proteins of the Ras superfamily. Each GTP-bound Rab protein resides in a particular type of endosome and functions by recruiting specific effector proteins. Following their internalization into early RAB5-containing endosomes, receptors can rapidly recycle back to the plasma membrane by a RAB4-dependent mechanism, traffic to the recycling compartment that contains RAB11A or remain in endosomes, which mature into multivesicular bodies (MVBs) and late endosomes. MVBs are defined by the presence of intraluminal vesicles (ILVs) that are formed in a process of inward membrane invagination involving ESCRT (endosomal sorting complex required for transport) complexes. Early-to-late endosome maturation involves the acquisition of RAB7 and the removal of endosomal components that are capable of, and necessary for, recycling. In the MVBs, cargo destined for degradation is incorporated into ILVs. Fusion of late endosomes and MVBs with lysosomes carrying proteolytic enzymes results in cargo degradation.
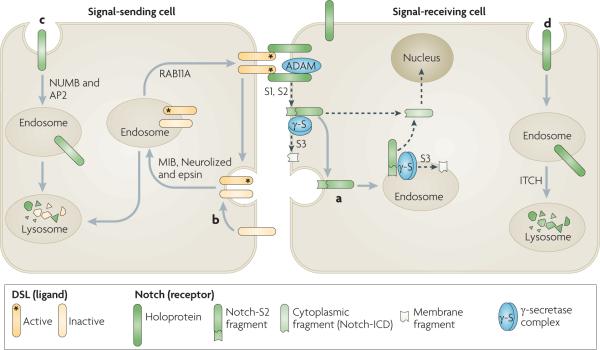
Dsl (Delta/Serrate/lag2 domain) proteins, which are Notch ligands, are transmembrane proteins that are present in the plasma membrane of a signal-sending cell. Dsl proteins interact with Notch located at the surface of a signal-receiving cell, which results in proteolytic S1 and S2 cleavages of the extracellular domain of Notch mediated by a disintegrin and metalloprotease (ADAM) enzyme and generates a Notch extracellular truncation fragment (Notch-S2) (see the figure, part a). Notch-S2 is further subject to intramembrane S3 cleavage by the γ-secretase complex, either in the plasma membrane or after endocytosis, to generate a membrane fragment and the cytoplasmic intracellular domain fragment (Notch-ICD), which translocates to the nucleus to activate the transcription of target genes. Evidence in Drosophila melanogaster and mammalian cells suggests that endocytosis is necessary for the S3 cleavage of recombinant and endogenous Notch,. Although γ-secretase is present in both the plasma membrane and endosomes,, it has optimal activity at the low pH levels of endosomes and lysosomes,,. The surface concentration of functional Dsl proteins is crucial for physiological levels of Notch signalling, and this is regulated by endocytosis (reviewed in REF. 134). The endocytosis and recycling of the Notch ligand Delta in signal-sending cells is also required for Delta activation, although the mechanism is not understood. Four E3 RING ubiquitin ligases, MIB1 and MIB2 and Neurolized 1 and Neurolized 2, as well as Epsin and RAB11A, are implicated in Dsl internalization and recycling (see the figure, part b)–. The endocytic adaptors NUMB and AP2 promote the endocytosis of non-ligand bound Notch and its degradation in the lysosome of the signal-sending cell (see the figure, part c). In the D. melanogaster pupal retina, Notch is endocytosed and directed to late endosomes and lysosomes in the signal-receiving cell (see the figure, part d)–. In mammalian cells targeting of non-ligand bound Notch to lysosomes is promoted by the lysine 29-linked polyubiquitylation of Notch, which is mediated by the HECT domain-containing E3 ligase ITCH (also known as AIP4).
References
-
- Polo S, Di Fiore PP. Endocytosis conducts the cell signaling orchestra. Cell. 2006;124:897–900. - PubMed
-
- Conner SD, Schmid SL. Regulated portals of entry into the cell. Nature. 2003;422:37–44. - PubMed
-
- Sorkin A. Cargo recognition during clathrin-mediated endocytosis: a team effort. Curr. Opin. Cell Biol. 2004;16:392–399. - PubMed
-
- Doherty GJ, McMahon HT. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009;78:857–902. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
