

Generation of new protein functions by nonhomologous combinations and rearrangements of domains and modules
- PMID: 19700302
- PMCID: PMC2763956
- DOI: 10.1016/j.copbio.2009.07.007
Generation of new protein functions by nonhomologous combinations and rearrangements of domains and modules
Abstract
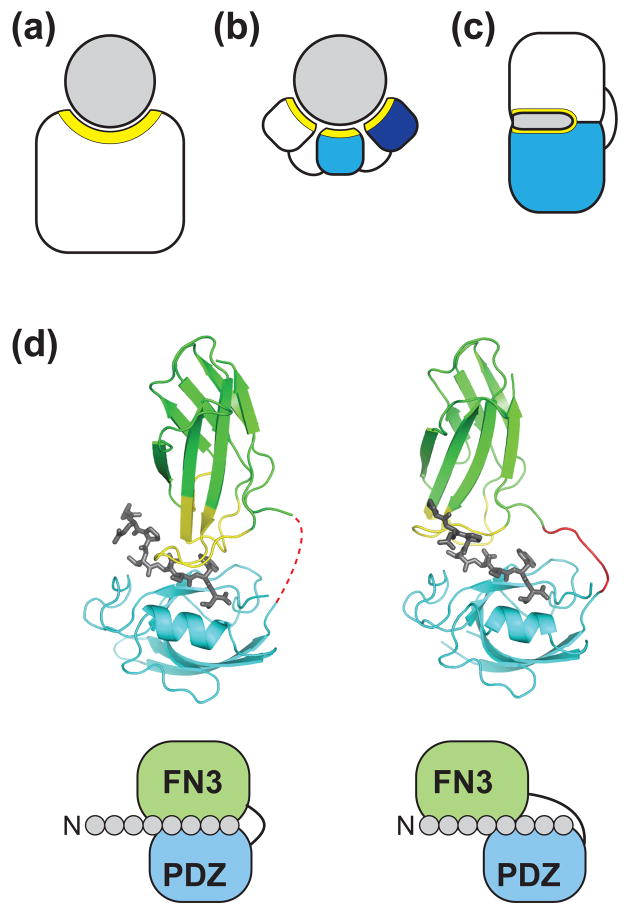
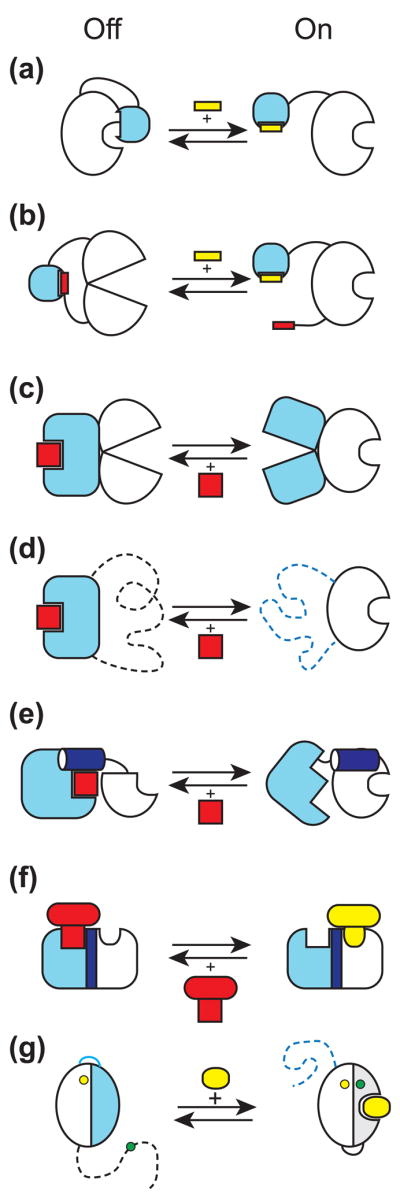
Generation of novel protein functions is a major goal in biotechnology and also a rigorous test for our understanding of the relationship between protein structure and function. Early examples of protein engineering focused on design and directed evolution within the constraints of the original protein architecture, exemplified by the highly successful fields of antibody and enzyme engineering. Recent studies show that protein engineering strategies which step away from these natural architectures, that is by manipulating the organization of domains and modules thus mimicking nonhomologous recombination, are highly effective in producing complex and sophisticated functions in terms of both molecular recognition and regulation.
Figures
References
-
- Pluckthun A, Mayo SL. The design of evolution and the evolution of design. Curr Opin Struct Biol. 2007;17:451–453.
-
- Kortemme T, Baker D. Computational design of protein-protein interactions. Curr Opin Chem Biol. 2004;8:91–97. - PubMed
-
- Winter G, Griffiths AD, Hawkins RE, Hoogenboom HR. Making antibodies by phage display technology. Annu Rev Immunol. 1994;12:433–455. - PubMed
-
- Binz HK, Amstutz P, Pluckthun A. Engineering novel binding proteins from nonimmunoglobulin domains. Nat Biotechnol. 2005;23:1257–1268. - PubMed
-
- Looger LL, Dwyer MA, Smith JJ, Hellinga HW. Computational design of receptor and sensor proteins with novel functions. Nature. 2003;423:185–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
