

At4g24160, a soluble acyl-coenzyme A-dependent lysophosphatidic acid acyltransferase
- PMID: 19700561
- PMCID: PMC2754629
- DOI: 10.1104/pp.109.144261
At4g24160, a soluble acyl-coenzyme A-dependent lysophosphatidic acid acyltransferase
Abstract
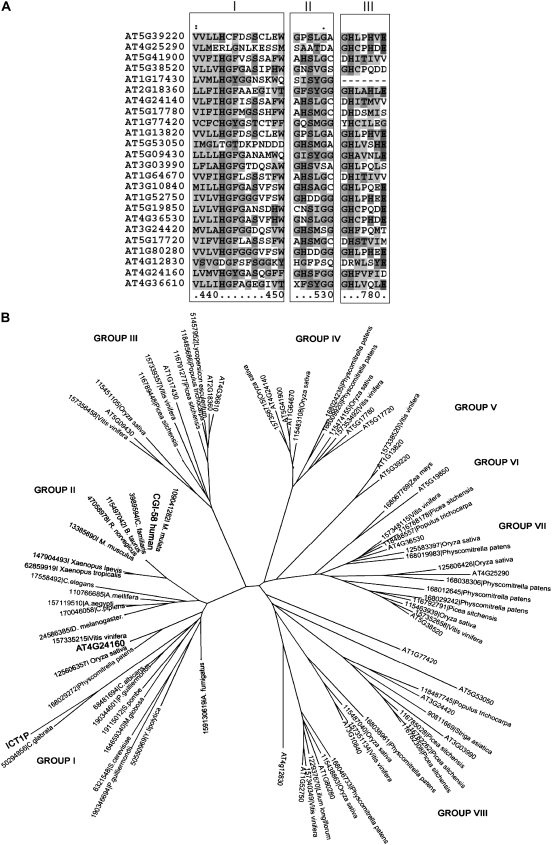
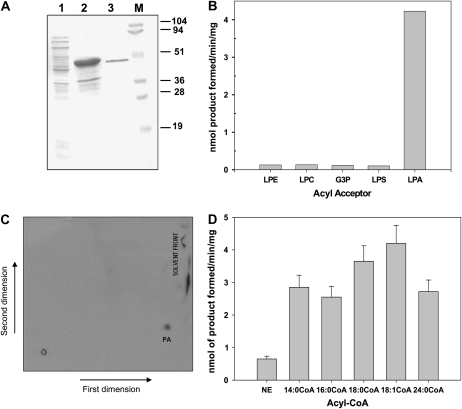
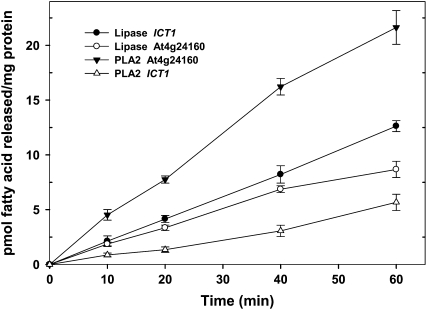
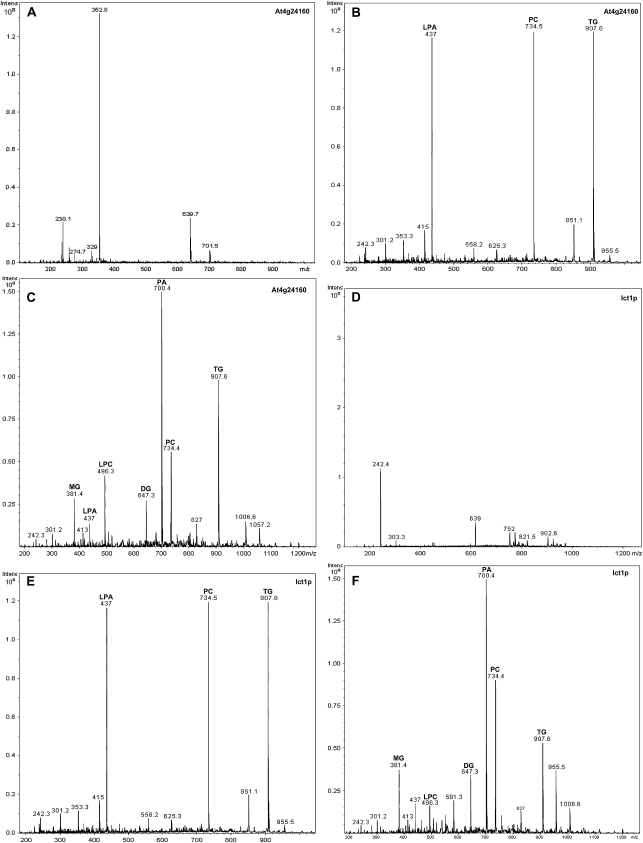
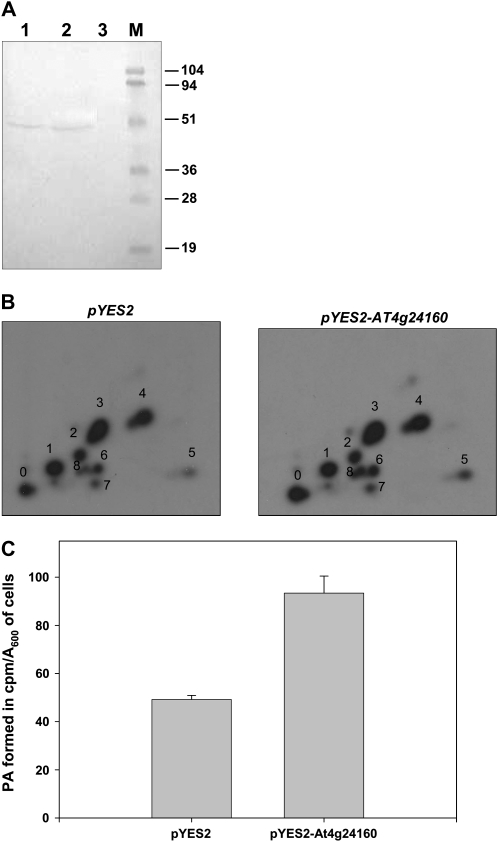
Human CGI-58 (for comparative gene identification-58) and YLR099c, encoding Ict1p in Saccharomyces cerevisiae, have recently been identified as acyl-CoA-dependent lysophosphatidic acid acyltransferases. Sequence database searches for CGI-58 like proteins in Arabidopsis (Arabidopsis thaliana) revealed 24 proteins with At4g24160, a member of the alpha/beta-hydrolase family of proteins being the closest homolog. At4g24160 contains three motifs that are conserved across the plant species: a GXSXG lipase motif, a HX(4)D acyltransferase motif, and V(X)(3)HGF, a probable lipid binding motif. Dendrogram analysis of yeast ICT1, CGI-58, and At4g24160 placed these three polypeptides in the same group. Here, we describe and characterize At4g24160 as, to our knowledge, the first soluble lysophosphatidic acid acyltransferase in plants. A lipidomics approach revealed that At4g24160 has additional triacylglycerol lipase and phosphatidylcholine hydrolyzing enzymatic activities. These data establish At4g24160, a protein with a previously unknown function, as an enzyme that might play a pivotal role in maintaining the lipid homeostasis in plants by regulating both phospholipid and neutral lipid levels.
Figures

References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
