

Sustained cyclic AMP production by parathyroid hormone receptor endocytosis
- PMID: 19701185
- PMCID: PMC3032084
- DOI: 10.1038/nchembio.206
Sustained cyclic AMP production by parathyroid hormone receptor endocytosis
Abstract
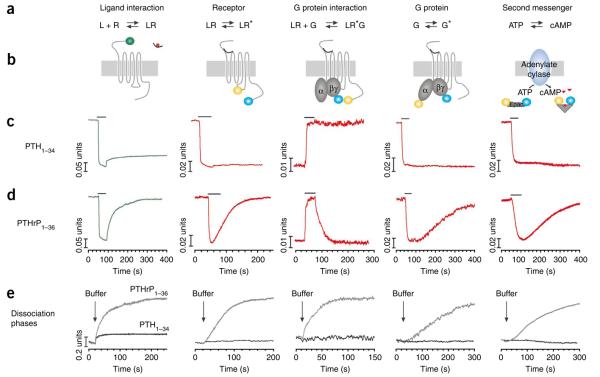
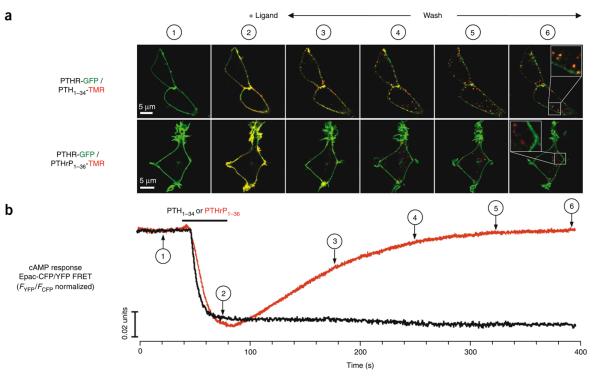
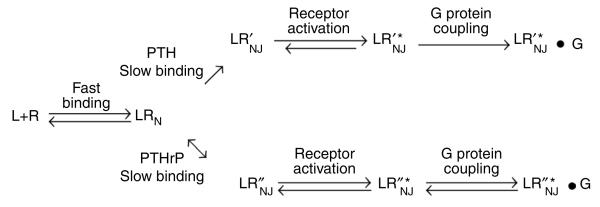
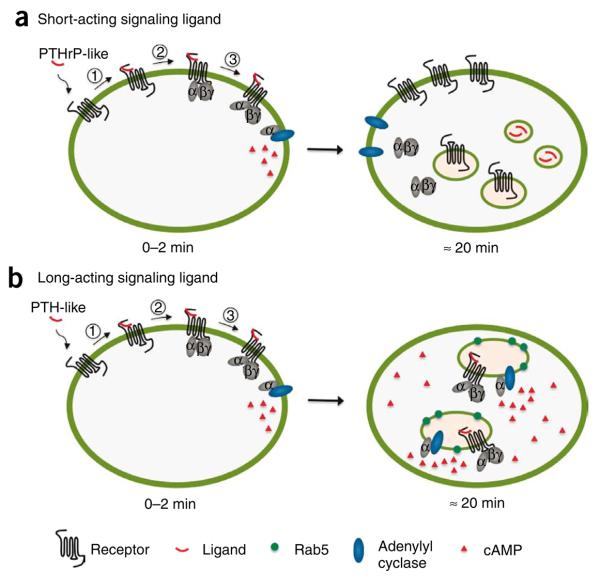
Cell signaling mediated by the G protein-coupled parathyroid hormone receptor type 1 (PTHR) is fundamental to bone and kidney physiology. It has been unclear how the two ligand systems--PTH, endocrine and homeostatic, and PTH-related peptide (PTHrP), paracrine--can effectively operate with only one receptor and trigger different durations of the cAMP responses. Here we analyze the ligand response by measuring the kinetics of activation and deactivation for each individual reaction step along the PTHR signaling cascade. We found that during the time frame of G protein coupling and cAMP production, PTHrP(1-36) action was restricted to the cell surface, whereas PTH(1-34) had moved to internalized compartments where it remained associated with the PTHR and Galpha(s), potentially as a persistent and active ternary complex. Such marked differences suggest a mechanism by which PTH and PTHrP induce differential responses, and these results indicate that the central tenet that cAMP production originates exclusively at the cell membrane must be revised.
Figures


Comment in
-
When two keys fit one lock, surprises follow.Nat Chem Biol. 2009 Oct;5(10):707-8. doi: 10.1038/nchembio.225. Nat Chem Biol. 2009. PMID: 19763099 No abstract available.
References
-
- Lohse MJ, et al. Optical techniques to analyze real-time activation and signaling of G-protein-coupled receptors. Trends Pharmacol. Sci. 2008;29:159–165. - PubMed
-
- Pierce KL, Premont RT, Lefkowitz RJ. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002;3:639–650. - PubMed
-
- Pugh EN, Jr., Lamb TD. Amplification and kinetics of the activation steps in phototransduction. Biochim. Biophys. Acta. 1993;1141:111–149. - PubMed
-
- Drake MT, Shenoy SK, Lefkowitz RJ. Trafficking of G protein-coupled receptors. Circ. Res. 2006;99:570–582. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
