

Primary cilia can both mediate and suppress Hedgehog pathway-dependent tumorigenesis
- PMID: 19701205
- PMCID: PMC2895420
- DOI: 10.1038/nm.2011
Primary cilia can both mediate and suppress Hedgehog pathway-dependent tumorigenesis
Abstract
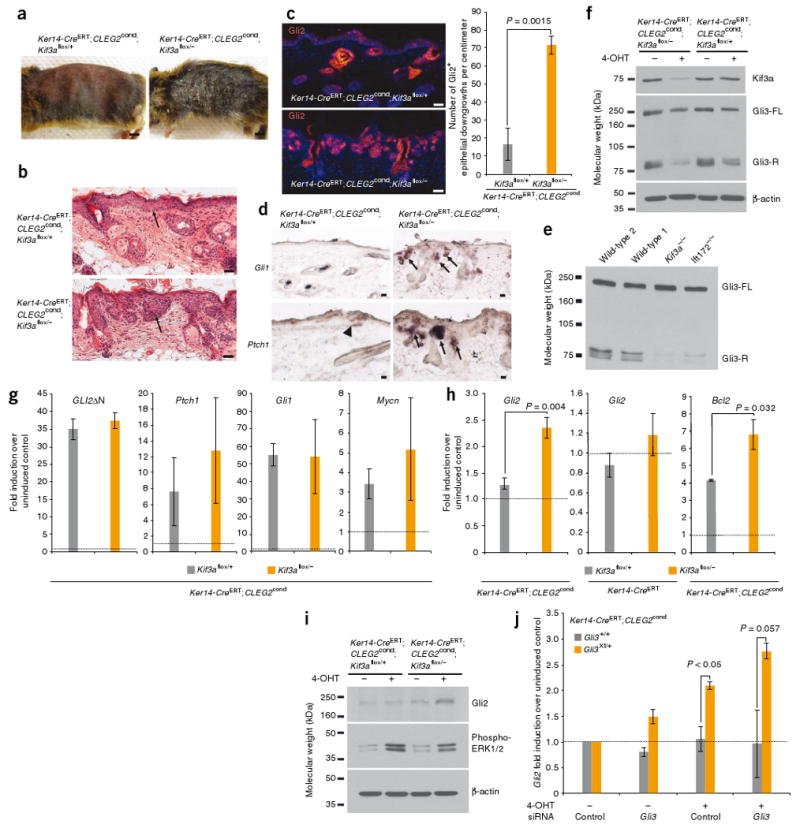
Primary cilia are present on most mammalian cells and are implicated in transducing Hedgehog (Hh) signals during development; however, the prevalence of cilia on human tumors remains unclear, and the role of cilia in cancer has not been examined. Here we show that human basal cell carcinomas (BCCs) are frequently ciliated, and we test the role of cilia in BCC by conditionally deleting Kif3a (encoding kinesin family member 3A) or Ift88 (encoding intraflagellar transport protein 88), genes required for ciliogenesis, in two Hh pathway-dependent mouse tumor models. Ciliary ablation strongly inhibited BCC-like tumors induced by an activated form of Smoothened. In contrast, removal of cilia accelerated tumors induced by activated Gli2, a transcriptional effector of Hh signaling. These seemingly paradoxical effects are consistent with a dual role for cilia in mediating both the activation and the repression of the Hh signaling pathway. Our findings demonstrate that cilia function as unique signaling organelles that can either mediate or suppress tumorigenesis depending on the nature of the oncogenic initiating event.
Figures



Comment in
-
Two sides to cilia in cancer.Nat Med. 2009 Sep;15(9):994-6. doi: 10.1038/nm0909-994. Nat Med. 2009. PMID: 19734870 No abstract available.
References
-
- Dahmane N, Lee J, Robins P, Heller P, Ruiz i Altaba A. Activation of the transcription factor Gli1 and the Sonic hedgehog signalling pathway in skin tumours. Nature. 1997;389:876–881. - PubMed
-
- Rubin LL, de Sauvage FJ. Targeting the Hedgehog pathway in cancer. Nat Rev Drug Discov. 2006;5:1026–1033. - PubMed
-
- Hooper JE, Scott MP. Communicating with Hedgehogs. Nat Rev Mol Cell Biol. 2005;6:306–317. - PubMed
-
- Xie J, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature. 1998;391:90–92. - PubMed
-
- Reifenberger J, et al. Missense mutations in SMOH in sporadic basal cell carcinomas of the skin and primitive neuroectodermal tumors of the central nervous system. Cancer Res. 1998;58:1798–1803. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
