

Altered Na+ transport after an intracellular alpha-subunit deletion reveals strict external sequential release of Na+ from the Na/K pump
- PMID: 19706387
- PMCID: PMC2741281
- DOI: 10.1073/pnas.0903752106
Altered Na+ transport after an intracellular alpha-subunit deletion reveals strict external sequential release of Na+ from the Na/K pump
Abstract
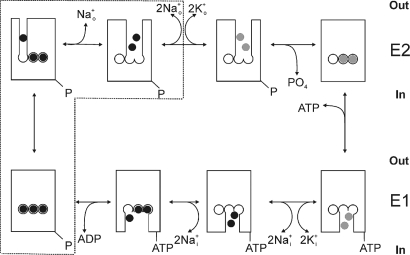
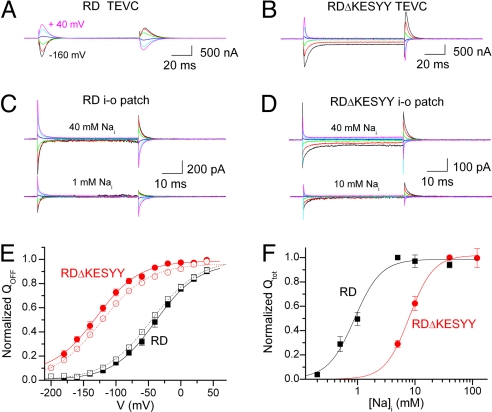
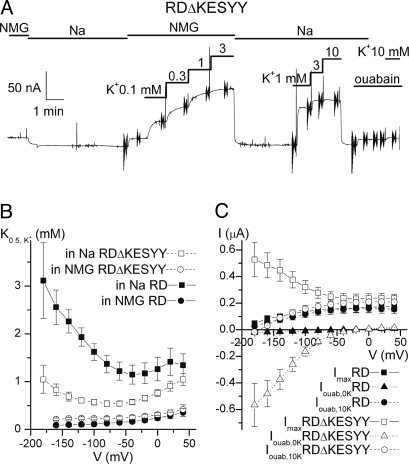
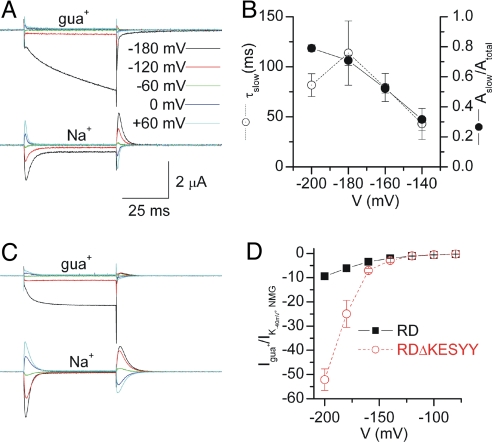
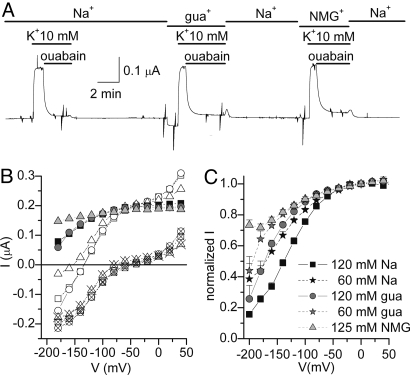
The Na/K pump actively exports 3 Na(+) in exchange for 2 K(+) across the plasmalemma of animal cells. As in other P-type ATPases, pump function is more effective when the relative affinity for transported ions is altered as the ion binding sites alternate between opposite sides of the membrane. Deletion of the five C-terminal residues from the alpha-subunit diminishes internal Na(+) (Na(i)(+)) affinity approximately 25-fold [Morth et al. (2007) Nature 450:1043-1049]. Because external Na(+) (Na(o)(+)) binding is voltage-dependent, we studied the reactions involving this process by using two-electrode and inside-out patch voltage clamp in normal and truncated (DeltaKESYY) Xenopus-alpha1 pumps expressed in oocytes. We observed that DeltaKESYY (i) decreased both Na(o)(+) and Na(i)(+) apparent affinities in the absence of K(o)(+), and (ii) did not affect apparent Na(o)(+) affinity at high K(o)(+). These results support a model of strict sequential external release of Na(+) ions, where the Na(+)-exclusive site releases Na(+) before the sites shared with K(+) and the DeltaKESYY deletion only reduces Na(o)(+) affinity at the shared sites. Moreover, at nonsaturating K(o)(+), DeltaKESYY induced an inward flow of Na(+) through Na/K pumps at negative potentials. Guanidinium(+) can also permeate truncated pumps, whereas N-methyl-D-glucamine cannot. Because guanidinium(o)(+) can also traverse normal Na/K pumps in the absence of both Na(o)(+) and K(o)(+) and can also inhibit Na/K pump currents in a Na(+)-like voltage-dependent manner, we conclude that the normal pathway transited by the first externally released Na(+) is large enough to accommodate guanidinium(+).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Post RL, Sen AK, Rosenthal AS. A phosphorylated intermediate in adenosine triphosphate-dependent sodium and potassium transport across kidney membranes. J Biol Chem. 1965;240:1437–1445. - PubMed
-
- Albers RW. Biochemical aspects of active transport. Annu Rev Biochem. 1967;36:727–756. - PubMed
-
- Morth JP, et al. Crystal structure of the sodium-potassium pump. Nature. 2007;450:1043–1049. - PubMed
-
- Geering K, et al. FXYD proteins: New tissue- and isoform-specific regulators of Na,K-ATPase. Ann NY Acad Sci. 2003;986:388–394. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
