

A luminal flavoprotein in endoplasmic reticulum-associated degradation
- PMID: 19706418
- PMCID: PMC2736413
- DOI: 10.1073/pnas.0900742106
A luminal flavoprotein in endoplasmic reticulum-associated degradation
Abstract
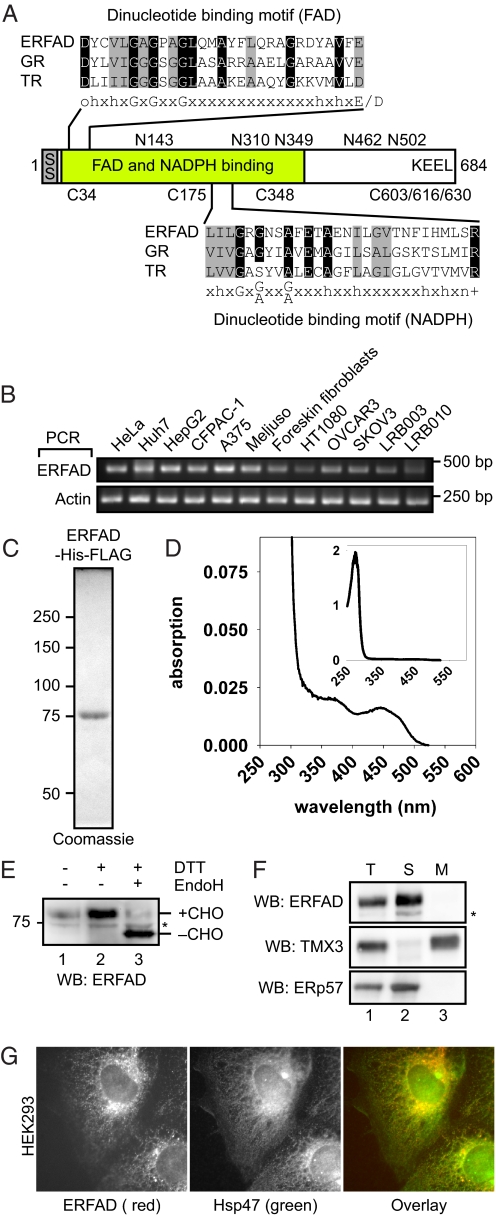
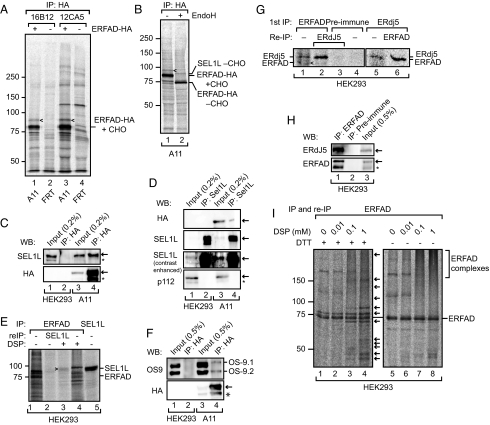
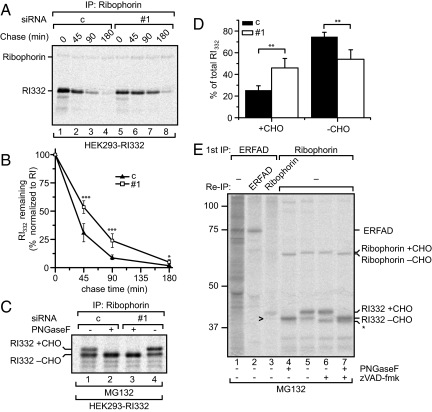
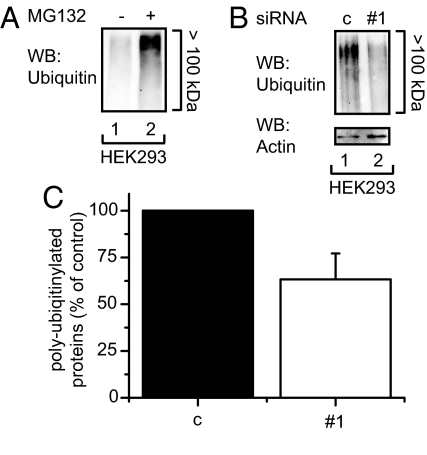
The quality control system of the endoplasmic reticulum (ER) discriminates between native and nonnative proteins. The latter are degraded by the ER-associated degradation (ERAD) pathway. Whereas many cytosolic and membrane components of this system are known, only few luminal players have been identified. In this study, we characterize ERFAD (ER flavoprotein associated with degradation), an ER luminal flavoprotein that functions in ERAD. Upon knockdown of ERFAD, the degradation of the ERAD model substrate ribophorin 332 is delayed, and the overall level of polyubiquitinated cellular proteins is decreased. We also identify the ERAD components SEL1L, OS-9 and ERdj5, a known reductase of ERAD substrates, as interaction partners of ERFAD. Our data show that ERFAD facilitates the dislocation of certain ERAD substrates to the cytosol, and we discuss the findings in relation to a potential redox function of the protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
