

Sparsification of neuronal activity in the visual cortex at eye-opening
- PMID: 19706480
- PMCID: PMC2736444
- DOI: 10.1073/pnas.0907660106
Sparsification of neuronal activity in the visual cortex at eye-opening
Abstract
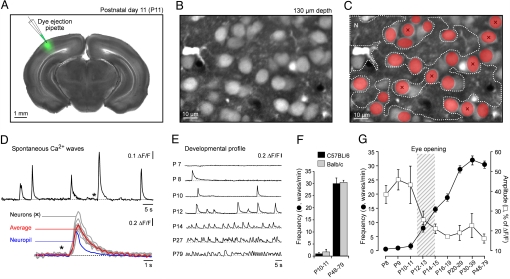
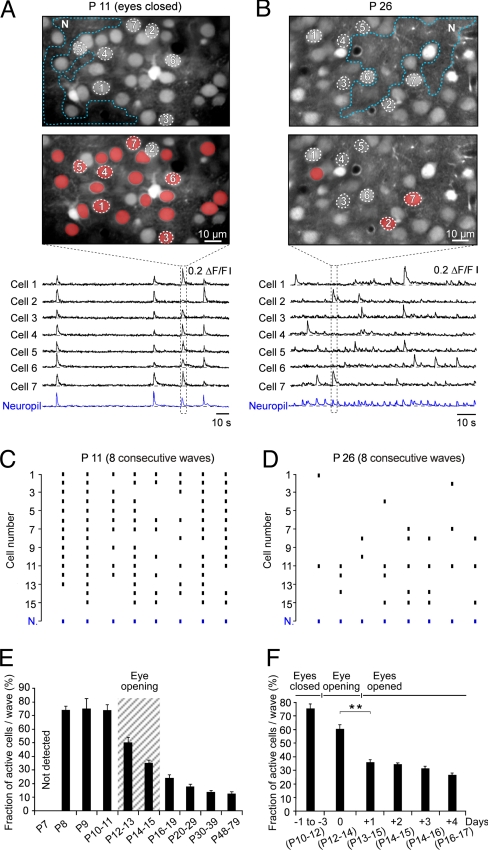
Eye-opening represents a turning point in the function of the visual cortex. Before eye-opening, the visual cortex is largely devoid of sensory inputs and neuronal activities are generated intrinsically. After eye-opening, the cortex starts to integrate visual information. Here we used in vivo two-photon calcium imaging to explore the developmental changes of the mouse visual cortex by analyzing the ongoing spontaneous activity. We found that before eye-opening, the activity of layer 2/3 neurons consists predominantly of slow wave oscillations. These waves were first detected at postnatal day 8 (P8). Their initial very low frequency (0.01 Hz) gradually increased during development to approximately 0.5 Hz in adults. Before eye-opening, a large fraction of neurons (>75%) was active during each wave. One day after eye-opening, this dense mode of recruitment changed to a sparse mode with only 36% of active neurons per wave. This was followed by a progressive decrease during the following weeks, reaching 12% of active neurons per wave in adults. The possible role of visual experience for this process of sparsification was investigated by analyzing dark-reared mice. We found that sparsification also occurred in these mice, but that the switch from a dense to a sparse activity pattern was delayed by 3-4 days as compared with normally-reared mice. These results reveal a modulatory contribution of visual experience during the first days after eye-opening, but an overall dominating role of intrinsic factors. We propose that the transformation in network activity from dense to sparse is a prerequisite for the changed cortical function at eye-opening.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Lopez-Bendito G, Molnar Z. Thalamocortical development: How are we going to get there? Nat Rev Neurosci. 2003;4:276–289. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
