

From the Cover: Osmotrophy in modular Ediacara organisms
- PMID: 19706530
- PMCID: PMC2732876
- DOI: 10.1073/pnas.0904836106
From the Cover: Osmotrophy in modular Ediacara organisms
Abstract
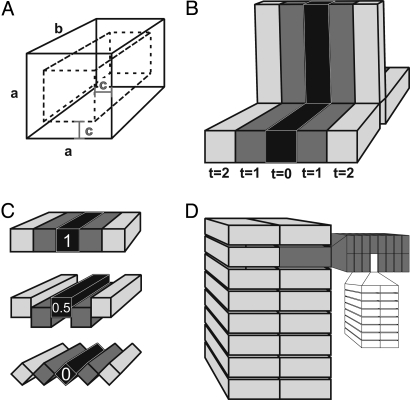
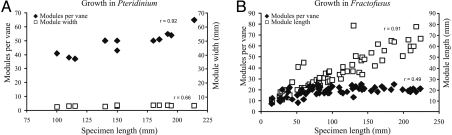
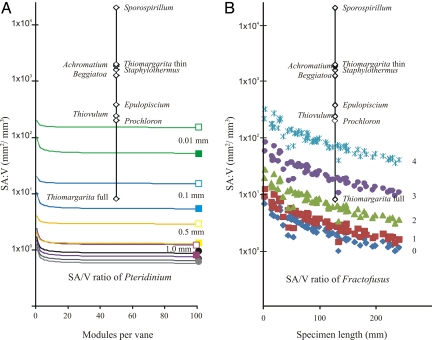
The Ediacara biota include macroscopic, morphologically complex soft-bodied organisms that appear globally in the late Ediacaran Period (575-542 Ma). The physiology, feeding strategies, and functional morphology of the modular Ediacara organisms (rangeomorphs and erniettomorphs) remain debated but are critical for understanding their ecology and phylogeny. Their modular construction triggered numerous hypotheses concerning their likely feeding strategies, ranging from micro-to-macrophagus feeding to photoautotrophy to osmotrophy. Macrophagus feeding in rangeomorphs and erniettomorphs is inconsistent with their lack of oral openings, and photoautotrophy in rangeomorphs is contradicted by their habitats below the photic zone. Here, we combine theoretical models and empirical data to evaluate the feasibility of osmotrophy, which requires high surface area to volume (SA/V) ratios, as a primary feeding strategy of rangeomorphs and erniettomorphs. Although exclusively osmotrophic feeding in modern ecosystems is restricted to microscopic bacteria, this study suggests that (i) fractal branching of rangeomorph modules resulted in SA/V ratios comparable to those observed in modern osmotrophic bacteria, and (ii) rangeomorphs, and particularly erniettomorphs, could have achieved osmotrophic SA/V ratios similar to bacteria, provided their bodies included metabolically inert material. Thus, specific morphological adaptations observed in rangeomorphs and erniettomorphs may have represented strategies for overcoming physiological constraints that typically make osmotrophy prohibitive for macroscopic life forms. These results support the viability of osmotrophic feeding in rangeomorphs and erniettomorphs, help explain their taphonomic peculiarities, and point to the possible importance of earliest macroorganisms for cycling dissolved organic carbon that may have been present in abundance during Ediacaran times.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Clapham ME, Narbonne GM. Ediacaran epifaunal tiering. Geology. 2002;30:627–630.
-
- Clapham ME, Narbonne GM, Gehling JG. Paleoecology of the oldest-known animal communities: Ediacaran assemblages at Mistaken Point, Newfoundland. Paleobiology. 2003;29:527–544.
-
- Bambach RK, Bush AM, Erwin DH. Autecology and the filling of ecospace: Key metazoan radiations. Palaeontology. 2007;50:1–22.
-
- Seilacher A. The nature of vendobionts. In: Vickers-Rich P, Komarower P, editors. The Rise and Fall of the Ediacaran Biota. London: Geological Society; 2007. pp. 387–397. Special Publication 286.
-
- Xiao S, Laflamme M. On the eve of animal radiation: Phylogeny, ecology and evolution of the Ediacara biota. Trends Ecol Evol. 2009;24:31–40. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
