

Behavioral tagging is a general mechanism of long-term memory formation
- PMID: 19706547
- PMCID: PMC2732837
- DOI: 10.1073/pnas.0907078106
Behavioral tagging is a general mechanism of long-term memory formation
Abstract
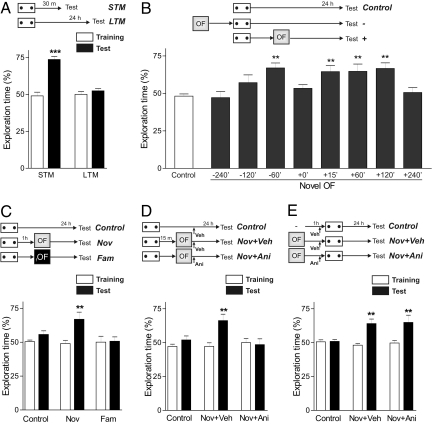
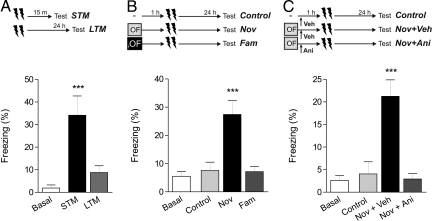
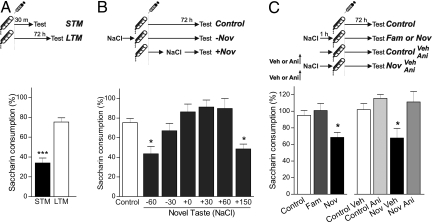
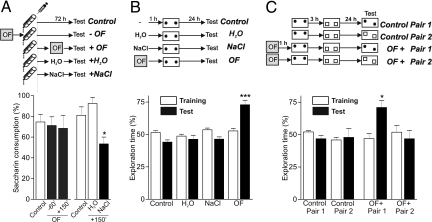
In daily life, memories are intertwined events. Little is known about the mechanisms involved in their interactions. Using two hippocampus-dependent (spatial object recognition and contextual fear conditioning) and one hippocampus-independent (conditioned taste aversion) learning tasks, we show that in rats subjected to weak training protocols that induce solely short term memory (STM), long term memory (LTM) is promoted and formed only if training sessions took place in contingence with a novel, but not familiar, experience occurring during a critical time window around training. This process requires newly synthesized proteins induced by novelty and reveals a general mechanism of LTM formation that begins with the setting of a "learning tag" established by a weak training. These findings represent the first comprehensive set of evidences indicating the existence of a behavioral tagging process that in analogy to the synaptic tagging and capture process, need the creation of a transient, protein synthesis-independent, and input specific tag.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Costa-Mattioli M, Sonenberg N. Translational control of gene expression: A molecular switch for memory storage. Prog Brain Res. 2008;169:81–95. - PubMed
-
- Hebb D, editor. The Organization of Behavior: A Neuropsychological Theory. New York: Wiley; 1949.
-
- Konorski J, editor. Conditioned Reflexes and Neuron Organization. Cambridge, UK: Cambridge Univ Press; 1948.
-
- Martin SJ, Grimwood PD, Morris RG. Synaptic plasticity and memory: An evaluation of the hypothesis. Annu Rev Neurosci. 2000;23:649–711. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
