

doi: 10.1182/blood-2009-07-231894.
Epub 2009 Aug 25.
Stage-specific susceptibility of human erythroblasts to Plasmodium falciparum malaria infection
Affiliations
- PMID: 19706885
- PMCID: PMC2766680
- DOI: 10.1182/blood-2009-07-231894
Item in Clipboard
Stage-specific susceptibility of human erythroblasts to Plasmodium falciparum malaria infection
Blood.
.
Abstract
Malaria parasites are known to invade and develop in erythrocytes and reticulocytes, but little is known about their infection of nucleated erythroid precursors. We used an in vitro cell system that progressed through basophilic, polychromatic, orthochromatic, and reticulocyte stages to mature erythrocytes. We show that orthochromatic cells are the earliest stages that may be invaded by Plasmodium falciparum, the causative agent of fatal human malaria. Susceptibility to invasion is distinct from intracellular survival and occurs at a time of extensive erythroid remodeling. Together these data suggest that the potential for complexity of host interactions involved in infection may be vastly greater than hitherto realized.
Figures
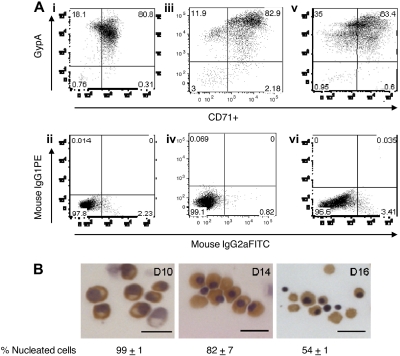
Differentiation of primary human erythroblasts in culture. CD34+-derived erythroid progenitors were differentiated into erythroblasts and reticulocytes after flow cytometry sorting for transferrin receptor (CD71)–positive cells. (A) Flow cytometry analysis for glycophorin A (gypA) and CD71 on days 10, 14, and 16 of culture. Day 10 (i), day 14 (iii), and day 16 (v) cells were at least 95% positive for erythroid cells. Panels ii, iv, and vi are isotype controls to show antibody specificity. (B) Hematoxylin/benzidine staining of day 10 polychromatic erythroblasts, day 14 early orthochromatic erythroblasts, and day 16 enucleating/reticulocytes. All populations contained hemoglobin (brown staining). Bar represents 20 μm.
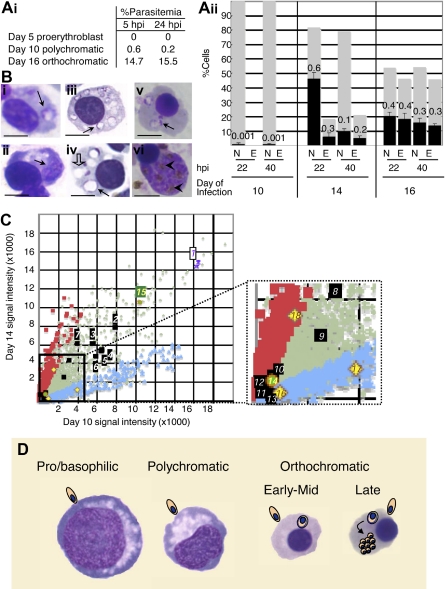
Characterization of human erythroblast maturation by susceptibility to Plasmodium falciparum malaria infection and by transcriptional response. (Ai) Percentage of day 5, 10, and 16 erythroblasts infected by Plasmodium falciparum. No infection is detected in pro-/basophilic erythroblasts (day 5). At day 10, when the population is largely at the polychromatic stage a few early orthochromatic cells are detected that can also be infected. (ii) Infection is seen in early/mid (day 14, ■) and late (day 16, ■) orthochromatic erythroblasts (N indicates nucleated; and E, enucleated), whereas only minor infection can be detected in polychromatic erythroblasts (day 10).  represent the percentage of nucleated/enucleated cells in the population, and ■ represent the proportion infected (marked with numbers) in each subset. Under these conditions, in cultures containing only mature erythrocytes we detect 60% infection. Error bars indicate SEM; n = 3 for day 10; n = 3 for day 14; n = 2 for day 16. (B) Brightfield images of Giemsa-stained infected erythroblast cells at 22 hours after invasion (hpi indicates hours of intracellular parasite growth; day 10, i; day 14, iii; day 16, v) and at 40 hpi (day 10, ii; day 14, iv; day 16, vi). On day 10 at 22 hpi, a few cells contain ring-stage parasites (i, ➙). Judging from staining of condensed nuclei, these cells are at the early orthochromatic stage. At 40 hpi, no mature schizonts are seen, but ring vacuoles (ii, ➙) may remain. On day 14, at 22 hpi, more than half the cells show rings (iii, ➙), which fail to mature to schizonts, but ring vacuoles (iv, ➙) are prominently detected. Half of these cells become vacuolated by 40 hpi (iv, [⇩]). On day 16, at 22 hpi and 40 hpi, respectively, ring-stage parasites (v, ➙) and schizont-stage parasites with hemozoin crystals (vi, ➤) are evident in nucleated, late orthochromatic erythroblasts. Photomicrographs were taken by light microscopy with a Zeiss Axioskop upright microscope and Nuance spectral camera/unmixing system (Cambridge Research and Instrumentation) using a 100× objective. Scale represents 5 μm. (C) Transcriptional profile of day 14 versus day 10 erythroblasts. Up-regulated transcripts are denoted with red squares; down-regulated, with blue diamonds; unchanged, with gray crosses. As expected, hemoglobins a and b were highly transcribed (spot 1, purple asterisks), erythrocyte protein band 4.1 (epb41) was up-regulated, and adducin 2 (add2) was down-regulated (inset, black squares, spots 12 and 13, respectively), suggesting assembly of the cytoskeleton was still in progress. Beta-actin (actb, spot 2), glycophorins (gypc spot 3, gypa spot 4, gypb spot 6), spectrin (spta1 spot 5, sptb spot 9), ankyrin (ank1 spot 7), band 3 (slc4a1 spot 8), Duffy (darc spot 10), and adducin 3 (add3 spot 11) are all expressed but show no significant change (fold change < 2, black squares). In addition erythropoietin receptor (epor spot 14, inset) transcript is reduced relative to transferrin receptor (cd71 spot 15, green circles) but nonetheless detectable. Bcl2-like 1 (bcl2l1 or bcl-xL; spot 18), a marker for terminal stages of erythropoiesis, is up-regulated 2.6-fold (P = .006, q = 0.005). In addition, it appears early orthochromatic cells are exiting the cell cycle because both cyclins D3 and E2 (ccnd3, ccne2; spots 16 and 17, respectively) are down-regulated ∼ 3-fold compared with polychromatic cells (P = .002, q = 0.003 and P = .02, q > 0.007, respectively). (D) Diagram displaying differential susceptibility of erythroblasts to P falciparum infection. Whereas orthochromatic cells can support parasite entry, only enucleating erythroblasts/nascent reticulocytes support intracellular parasite growth.
represent the percentage of nucleated/enucleated cells in the population, and ■ represent the proportion infected (marked with numbers) in each subset. Under these conditions, in cultures containing only mature erythrocytes we detect 60% infection. Error bars indicate SEM; n = 3 for day 10; n = 3 for day 14; n = 2 for day 16. (B) Brightfield images of Giemsa-stained infected erythroblast cells at 22 hours after invasion (hpi indicates hours of intracellular parasite growth; day 10, i; day 14, iii; day 16, v) and at 40 hpi (day 10, ii; day 14, iv; day 16, vi). On day 10 at 22 hpi, a few cells contain ring-stage parasites (i, ➙). Judging from staining of condensed nuclei, these cells are at the early orthochromatic stage. At 40 hpi, no mature schizonts are seen, but ring vacuoles (ii, ➙) may remain. On day 14, at 22 hpi, more than half the cells show rings (iii, ➙), which fail to mature to schizonts, but ring vacuoles (iv, ➙) are prominently detected. Half of these cells become vacuolated by 40 hpi (iv, [⇩]). On day 16, at 22 hpi and 40 hpi, respectively, ring-stage parasites (v, ➙) and schizont-stage parasites with hemozoin crystals (vi, ➤) are evident in nucleated, late orthochromatic erythroblasts. Photomicrographs were taken by light microscopy with a Zeiss Axioskop upright microscope and Nuance spectral camera/unmixing system (Cambridge Research and Instrumentation) using a 100× objective. Scale represents 5 μm. (C) Transcriptional profile of day 14 versus day 10 erythroblasts. Up-regulated transcripts are denoted with red squares; down-regulated, with blue diamonds; unchanged, with gray crosses. As expected, hemoglobins a and b were highly transcribed (spot 1, purple asterisks), erythrocyte protein band 4.1 (epb41) was up-regulated, and adducin 2 (add2) was down-regulated (inset, black squares, spots 12 and 13, respectively), suggesting assembly of the cytoskeleton was still in progress. Beta-actin (actb, spot 2), glycophorins (gypc spot 3, gypa spot 4, gypb spot 6), spectrin (spta1 spot 5, sptb spot 9), ankyrin (ank1 spot 7), band 3 (slc4a1 spot 8), Duffy (darc spot 10), and adducin 3 (add3 spot 11) are all expressed but show no significant change (fold change < 2, black squares). In addition erythropoietin receptor (epor spot 14, inset) transcript is reduced relative to transferrin receptor (cd71 spot 15, green circles) but nonetheless detectable. Bcl2-like 1 (bcl2l1 or bcl-xL; spot 18), a marker for terminal stages of erythropoiesis, is up-regulated 2.6-fold (P = .006, q = 0.005). In addition, it appears early orthochromatic cells are exiting the cell cycle because both cyclins D3 and E2 (ccnd3, ccne2; spots 16 and 17, respectively) are down-regulated ∼ 3-fold compared with polychromatic cells (P = .002, q = 0.003 and P = .02, q > 0.007, respectively). (D) Diagram displaying differential susceptibility of erythroblasts to P falciparum infection. Whereas orthochromatic cells can support parasite entry, only enucleating erythroblasts/nascent reticulocytes support intracellular parasite growth.
represent the percentage of nucleated/enucleated cells in the population, and ■ represent the proportion infected (marked with numbers) in each subset. Under these conditions, in cultures containing only mature erythrocytes we detect 60% infection. Error bars indicate SEM; n = 3 for day 10; n = 3 for day 14; n = 2 for day 16. (B) Brightfield images of Giemsa-stained infected erythroblast cells at 22 hours after invasion (hpi indicates hours of intracellular parasite growth; day 10, i; day 14, iii; day 16, v) and at 40 hpi (day 10, ii; day 14, iv; day 16, vi). On day 10 at 22 hpi, a few cells contain ring-stage parasites (i, ➙). Judging from staining of condensed nuclei, these cells are at the early orthochromatic stage. At 40 hpi, no mature schizonts are seen, but ring vacuoles (ii, ➙) may remain. On day 14, at 22 hpi, more than half the cells show rings (iii, ➙), which fail to mature to schizonts, but ring vacuoles (iv, ➙) are prominently detected. Half of these cells become vacuolated by 40 hpi (iv, [⇩]). On day 16, at 22 hpi and 40 hpi, respectively, ring-stage parasites (v, ➙) and schizont-stage parasites with hemozoin crystals (vi, ➤) are evident in nucleated, late orthochromatic erythroblasts. Photomicrographs were taken by light microscopy with a Zeiss Axioskop upright microscope and Nuance spectral camera/unmixing system (Cambridge Research and Instrumentation) using a 100× objective. Scale represents 5 μm. (C) Transcriptional profile of day 14 versus day 10 erythroblasts. Up-regulated transcripts are denoted with red squares; down-regulated, with blue diamonds; unchanged, with gray crosses. As expected, hemoglobins a and b were highly transcribed (spot 1, purple asterisks), erythrocyte protein band 4.1 (epb41) was up-regulated, and adducin 2 (add2) was down-regulated (inset, black squares, spots 12 and 13, respectively), suggesting assembly of the cytoskeleton was still in progress. Beta-actin (actb, spot 2), glycophorins (gypc spot 3, gypa spot 4, gypb spot 6), spectrin (spta1 spot 5, sptb spot 9), ankyrin (ank1 spot 7), band 3 (slc4a1 spot 8), Duffy (darc spot 10), and adducin 3 (add3 spot 11) are all expressed but show no significant change (fold change < 2, black squares). In addition erythropoietin receptor (epor spot 14, inset) transcript is reduced relative to transferrin receptor (cd71 spot 15, green circles) but nonetheless detectable. Bcl2-like 1 (bcl2l1 or bcl-xL; spot 18), a marker for terminal stages of erythropoiesis, is up-regulated 2.6-fold (P = .006, q = 0.005). In addition, it appears early orthochromatic cells are exiting the cell cycle because both cyclins D3 and E2 (ccnd3, ccne2; spots 16 and 17, respectively) are down-regulated ∼ 3-fold compared with polychromatic cells (P = .002, q = 0.003 and P = .02, q > 0.007, respectively). (D) Diagram displaying differential susceptibility of erythroblasts to P falciparum infection. Whereas orthochromatic cells can support parasite entry, only enucleating erythroblasts/nascent reticulocytes support intracellular parasite growth.References
-
- Panichakul T, Sattabongkot J, Chotivanich K, Sirichaisinthop J, Cui L, Udomsangpetch R. Production of erythropoietic cells in vitro for continuous culture of Plasmodium vivax. Int J Parasitol. 2007;37(14):1551–1557. - PubMed
-
- Pasvol G, Jungery M. Glycophorins and red cell invasion by Plasmodium falciparum. Ciba Found Symp. 1983;94:174–195. - PubMed
-
- Weatherall DJ. Genetic variation and susceptibility to infection: the red cell and malaria. Br J Haematol. 2008;141(3):276–286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI 039071/AI/NIAID NIH HHS/United States
- HL069630/HL/NHLBI NIH HHS/United States
- R01 CA098550/CA/NCI NIH HHS/United States
- T32 AI007476/AI/NIAID NIH HHS/United States
- R01CA98550/CA/NCI NIH HHS/United States
- R01 HL069630/HL/NHLBI NIH HHS/United States
- P01 HL 078826/HL/NHLBI NIH HHS/United States
- R01 HL 079397/HL/NHLBI NIH HHS/United States
- R01 AI039071/AI/NIAID NIH HHS/United States
- R01 HL079397/HL/NHLBI NIH HHS/United States
- P01 HL078826/HL/NHLBI NIH HHS/United States
- F30 HL094042/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
