

The CCR4-NOT complex physically and functionally interacts with TRAMP and the nuclear exosome
- PMID: 19707589
- PMCID: PMC2727002
- DOI: 10.1371/journal.pone.0006760
The CCR4-NOT complex physically and functionally interacts with TRAMP and the nuclear exosome
Abstract
Background: Ccr4-Not is a highly conserved multi-protein complex consisting in yeast of 9 subunits, including Not5 and the major yeast deadenylase Ccr4. It has been connected functionally in the nucleus to transcription by RNA polymerase II and in the cytoplasm to mRNA degradation. However, there has been no evidence so far that this complex is important for RNA degradation in the nucleus.
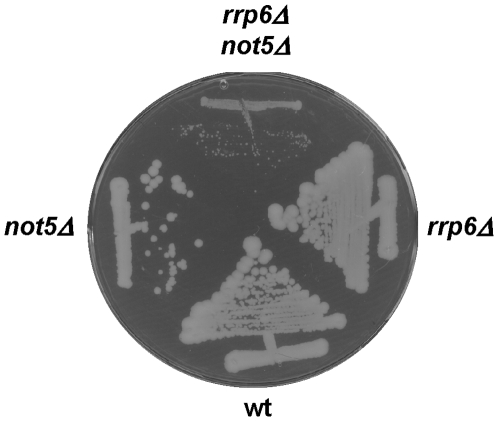
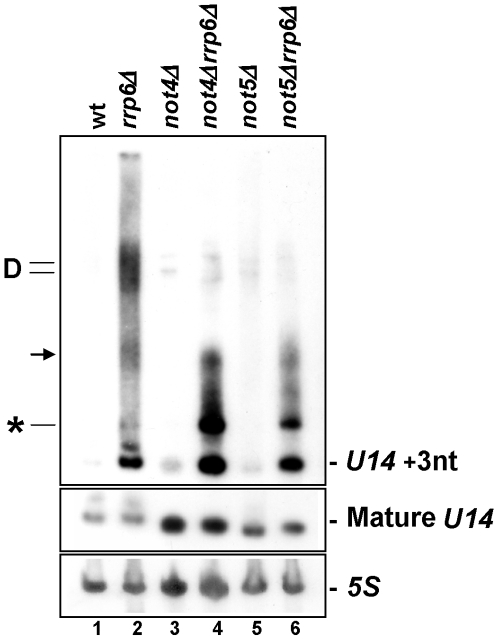
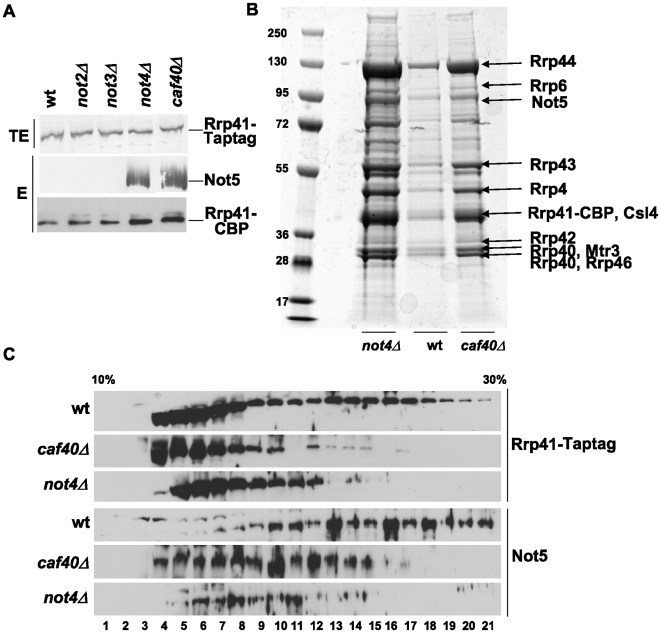
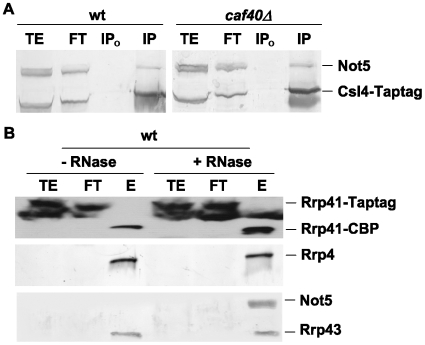
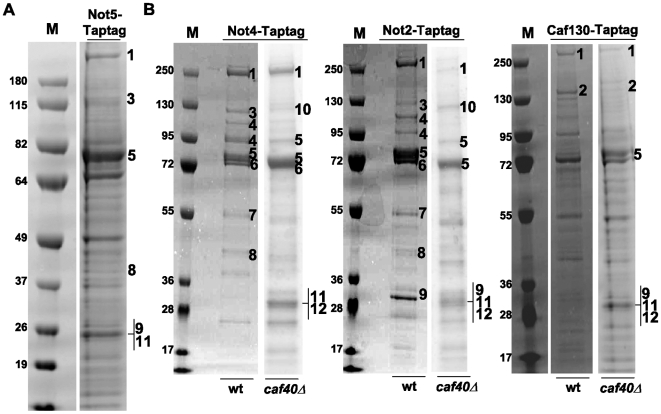
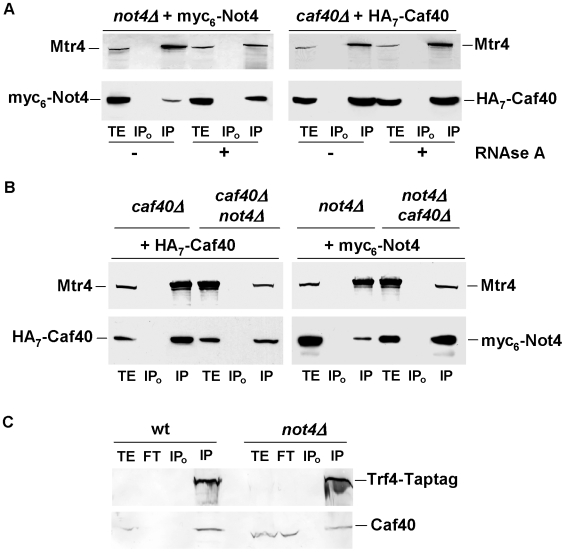
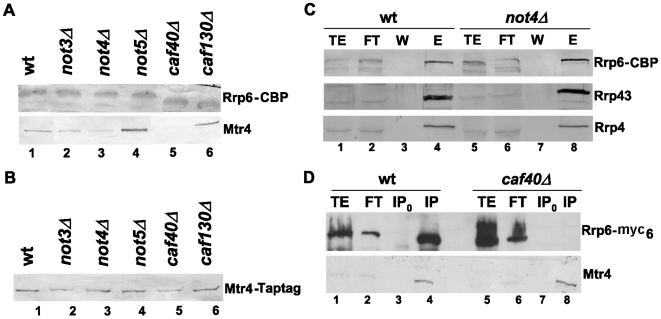
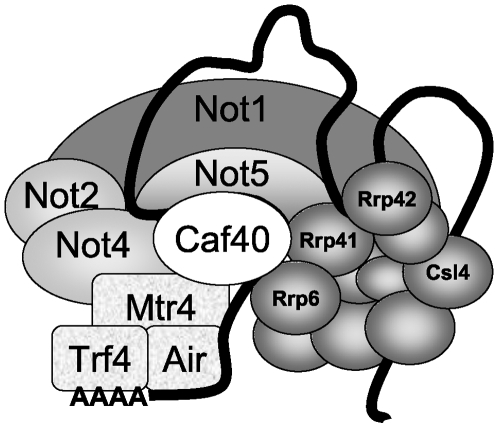
Methodology/principal findings: In this work we point to a new role for the Ccr4-Not complex in nuclear RNA metabolism. We determine the importance of the Ccr4-Not complex for the levels of non-coding nuclear RNAs, such as mis-processed and polyadenylated snoRNAs, whose turnover depends upon the nuclear exosome and TRAMP. Consistently, mutation of both the Ccr4-Not complex and the nuclear exosome results in synthetic slow growth phenotypes. We demonstrate physical interactions between the Ccr4-Not complex and the exosome. First, Not5 co-purifies with the exosome. Second, several exosome subunits co-purify with the Ccr4-Not complex. Third, the Ccr4-Not complex is important for the integrity of large exosome-containing complexes. Finally, we reveal a connection between the Ccr4-Not complex and TRAMP through the association of the Mtr4 helicase with the Ccr4-Not complex and the importance of specific subunits of Ccr4-Not for the association of Mtr4 with the nuclear exosome subunit Rrp6.
Conclusions/significance: We propose a model in which the Ccr4-Not complex may provide a platform contributing to dynamic interactions between the nuclear exosome and its co-factor TRAMP. Our findings connect for the first time the different players involved in nuclear and cytoplasmic RNA degradation.
Conflict of interest statement
Figures

References
-
- Butler JS. The yin and yang of the exosome. Trends Cell Biol. 2002;12:90–96. - PubMed
-
- Liu Q, Greimann JC, Lima CD. Reconstitution, activities, and structure of the eukaryotic RNA exosome. Cell. 2006;127:1223–1237. - PubMed
-
- Dziembowski A, Lorentzen E, Conti E, Seraphin B. A single subunit, Dis3, is essentially responsible for yeast exosome core activity. Nat Struct Mol Biol. 2007;14:15–22. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
