

BRCA2 interacts with the cytoskeletal linker protein plectin to form a complex controlling centrosome localization
- PMID: 19709076
- PMCID: PMC11158164
- DOI: 10.1111/j.1349-7006.2009.01282.x
BRCA2 interacts with the cytoskeletal linker protein plectin to form a complex controlling centrosome localization
Abstract
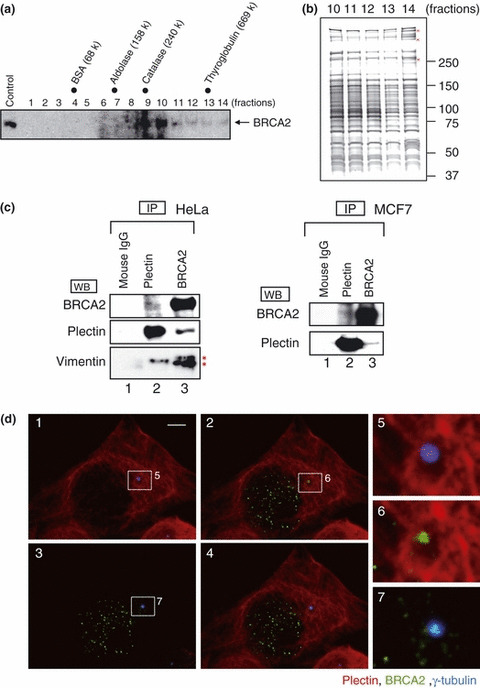
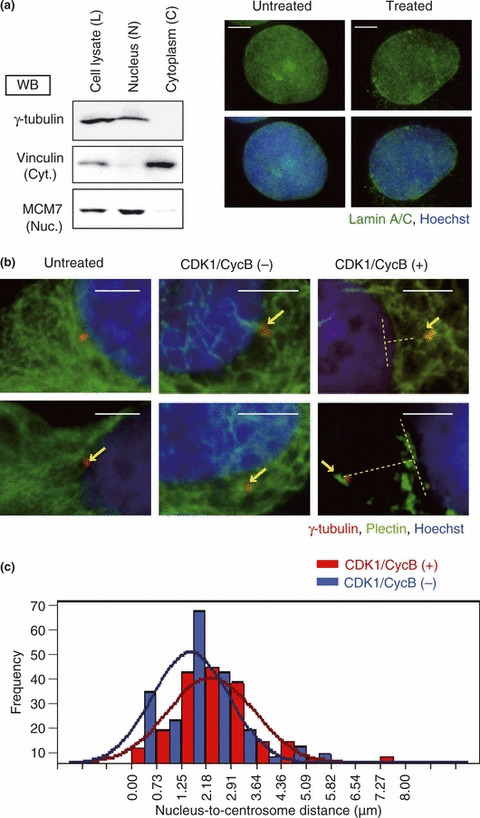
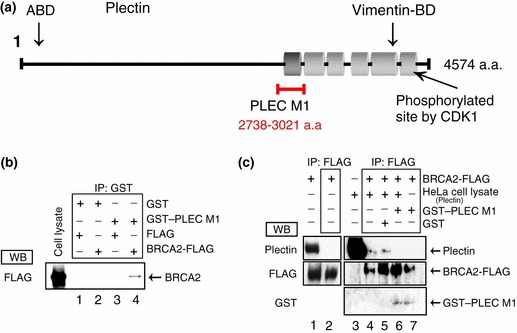
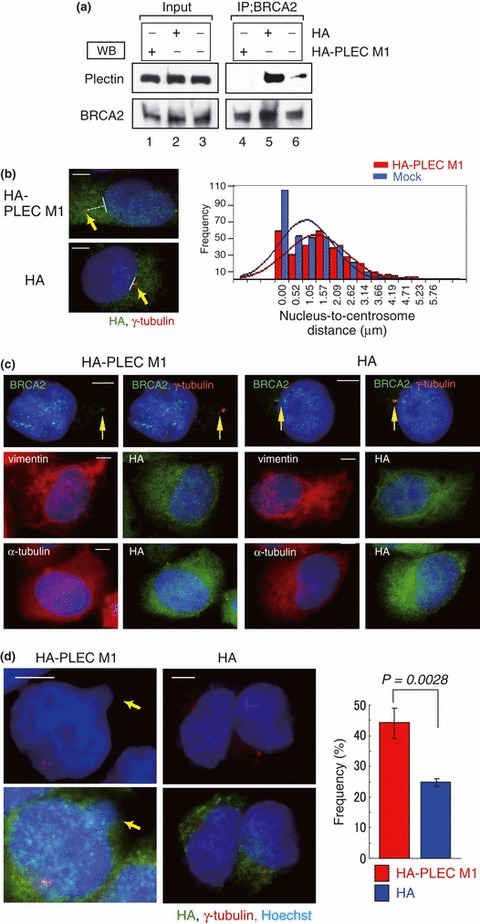
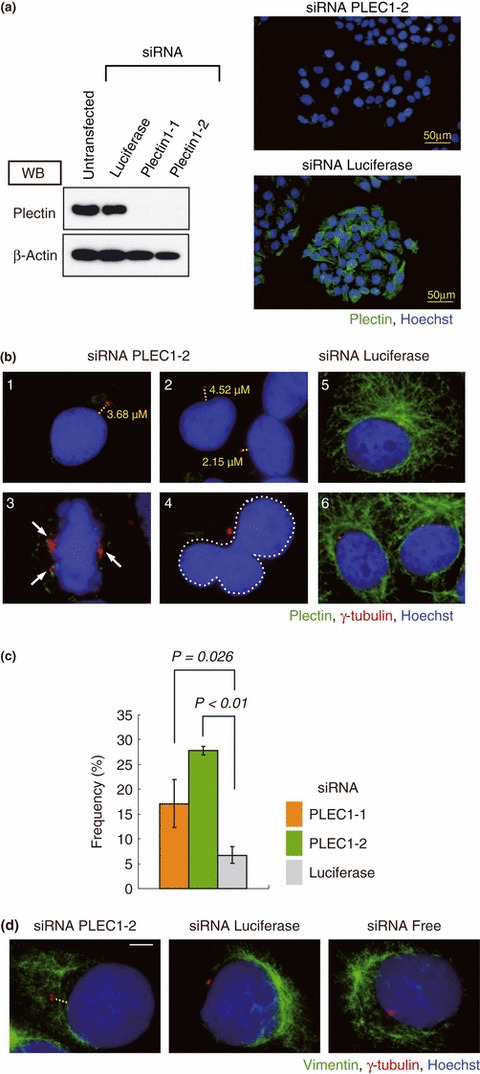
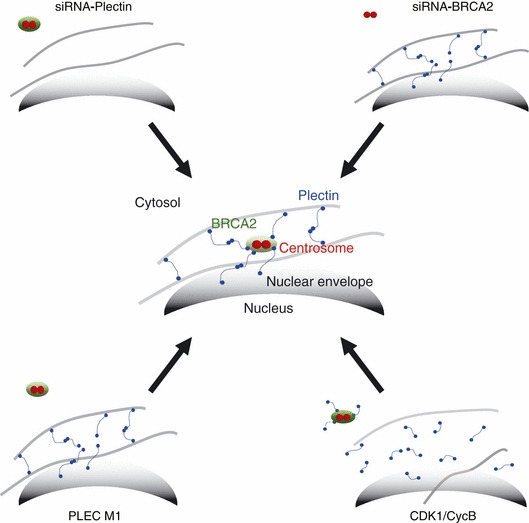
The breast cancer susceptibility gene (BRCA2) is localized mainly in the nucleus where it plays an important role in DNA damage repair. Some BRCA2 protein is also present in the centrosome. Here, we demonstrate that BRCA2 interacts with plectin, a cytoskeletal cross-linker protein, and that this interaction controls the position of the centrosome. Phosphorylation of plectin by cyclin-dependent kinase 1/cyclin B (CDK1/CycB) kinase has been reported to abolish its cross-linking function during mitosis. Here, we induced phosphorylation of plectin in prepared fractions of HeLa cells by adding activated CDK1/CycB kinase. Consequently, there was significant dissociation of the centrosome from the nuclear membrane. Plectin has six homologous ankyrin-like repeat domains (termed PLEC M1-M6). Using a pull-down assay, we found that GST-PLEC M1 and a GST-C-terminal region fusion protein (which comprised PLEC M6, along with an adjacent vimentin site) interacted with BRCA2. Since each PLEC module exhibits high homology to the others, the possibility of all six domains participating in this interaction was indicated. Moreover, when PLEC M1 was overexpressed in HeLa cells, it competed with endogenous plectin and inhibited the BRCA2-plectin interaction. This inhibitory effect resulted in dissociation of the centrosomes from the nucleus and increased the rate of micronuclei formation which may lead to carcinogenesis. In addition, when either BRCA2 or plectin was suppressed by the appropriate siRNA, a similar change in centrosomal positioning was observed. We suggest that the BRCA2-plectin interaction plays an important role in the regulation of centrosome localization and also that displacement of the centrosome may result in genomic instability and cancer development.
Figures
References
-
- Wooster R, Bignell G, Lancaster J et al. Identification of the breast cancer susceptibility gene BRCA2. Nature 1995; 378: 789–92. - PubMed
-
- Venkitaraman A. Cancer susceptibility and the functions of BRCA1 and BRCA2. Cell 2002; 108: 171–82. - PubMed
-
- Pellegrini L, Venkitaraman A. Emerging functions of BRCA2 in DNA recombination. Trends Biochem Sci 2004; 29: 310–6. - PubMed
-
- Esashi F, Christ N, Gannon J et al. CDK‐dependent phosphorylation of BRCA2 as a regulatory mechanism for recombinational repair. Nature 2005; 434: 598–604. - PubMed
-
- Wong A, Pero R, Ormonde P, Tavtigian S, Bartel P. RAD51 interacts with the evolutionarily conserved BRC motifs in the human breast cancer susceptibility gene brca2. J Biol Chem 1997; 272: 31941–4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
