

When worlds collide: IP(3) receptors and the ERAD pathway
- PMID: 19709743
- PMCID: PMC2752845
- DOI: 10.1016/j.ceca.2009.05.002
When worlds collide: IP(3) receptors and the ERAD pathway
Abstract
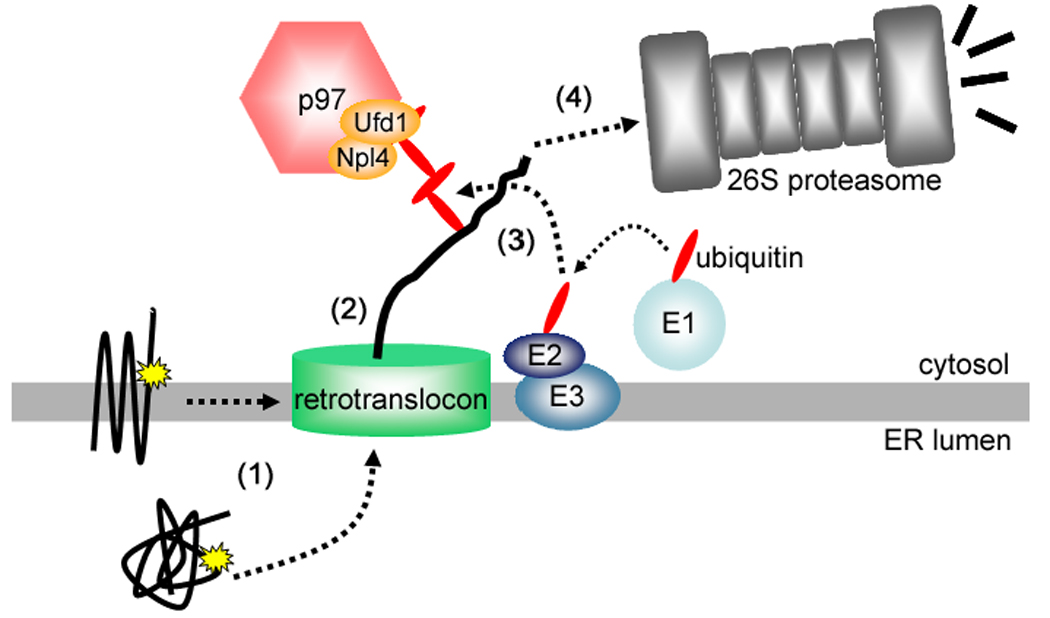
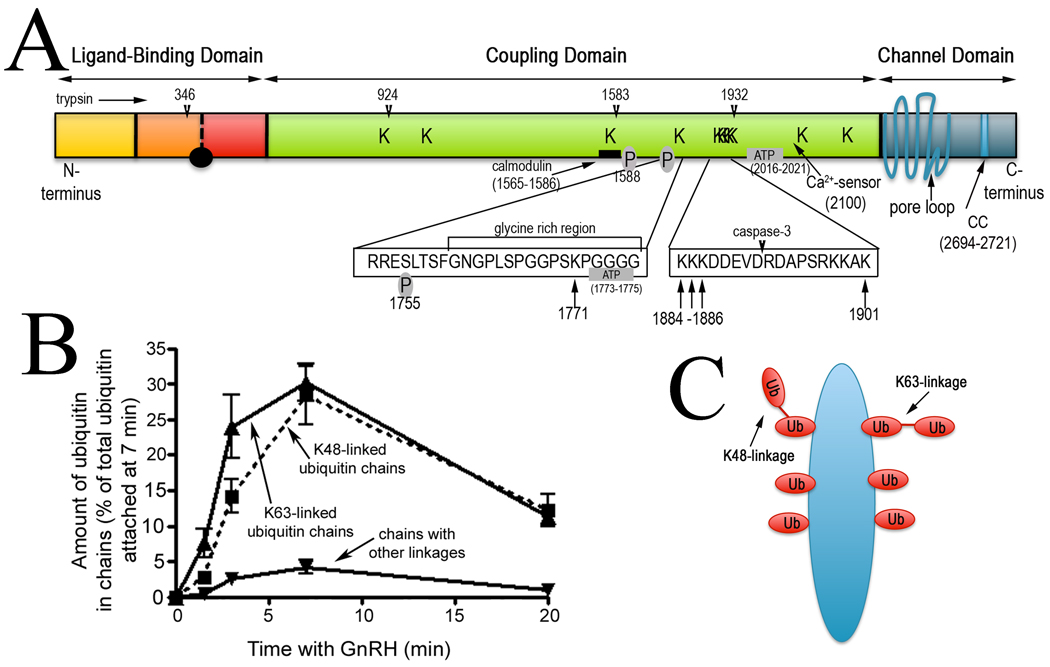
While cell signaling devotees tend to think of the endoplasmic reticulum (ER) as a Ca(2+) store, those who study protein synthesis tend to see it more as site for protein maturation, or even degradation when proteins do not fold properly. These two worldviews collide when inositol 1,4,5-trisphosphate (IP(3)) receptors are activated, since in addition to acting as release channels for stored ER Ca(2+), IP(3) receptors are rapidly destroyed via the ER-associated degradation (ERAD) pathway, a ubiquitination- and proteasome-dependent mechanism that clears the ER of aberrant proteins. Here we review recent studies showing that activated IP(3) receptors are ubiquitinated in an unexpectedly complex manner, and that a novel complex composed of the ER membrane proteins SPFH1 and SPFH2 (erlin 1 and 2) binds to IP(3) receptors immediately after they are activated and mediates their ERAD. Remarkably, it seems that the conformational changes that underpin channel opening make IP(3) receptors resemble aberrant proteins, which triggers their binding to the SPFH1/2 complex, their ubiquitination and extraction from the ER membrane and finally, their degradation by the proteasome. This degradation of activated IP(3) receptors by the ERAD pathway serves to reduce the sensitivity of ER Ca(2+) stores to IP(3) and may protect cells against deleterious effects of over-activation of Ca(2+) signaling pathways.
Figures


Similar articles
-
SPFH1 and SPFH2 mediate the ubiquitination and degradation of inositol 1,4,5-trisphosphate receptors in muscarinic receptor-expressing HeLa cells.Biochim Biophys Acta. 2009 Nov;1793(11):1710-8. doi: 10.1016/j.bbamcr.2009.09.004. Epub 2009 Sep 12. Biochim Biophys Acta. 2009. PMID: 19751772 Free PMC article.
-
An endoplasmic reticulum (ER) membrane complex composed of SPFH1 and SPFH2 mediates the ER-associated degradation of inositol 1,4,5-trisphosphate receptors.J Biol Chem. 2009 Apr 17;284(16):10433-45. doi: 10.1074/jbc.M809801200. Epub 2009 Feb 24. J Biol Chem. 2009. PMID: 19240031 Free PMC article.
-
SPFH2 mediates the endoplasmic reticulum-associated degradation of inositol 1,4,5-trisphosphate receptors and other substrates in mammalian cells.J Biol Chem. 2007 Jul 13;282(28):20104-15. doi: 10.1074/jbc.M701862200. Epub 2007 May 14. J Biol Chem. 2007. PMID: 17502376
-
Membrane Protein Quantity Control at the Endoplasmic Reticulum.J Membr Biol. 2017 Aug;250(4):379-392. doi: 10.1007/s00232-016-9931-0. Epub 2016 Oct 14. J Membr Biol. 2017. PMID: 27743014 Free PMC article. Review.
-
Chapter 4 - Inositol 1,4,5-Trisphosphate Receptor Ubiquitination.Prog Mol Biol Transl Sci. 2016;141:141-59. doi: 10.1016/bs.pmbts.2016.02.004. Epub 2016 Mar 30. Prog Mol Biol Transl Sci. 2016. PMID: 27378757 Review.
Cited by
-
Bcl-2-Protein Family as Modulators of IP3 Receptors and Other Organellar Ca2+ Channels.Cold Spring Harb Perspect Biol. 2020 Apr 1;12(4):a035089. doi: 10.1101/cshperspect.a035089. Cold Spring Harb Perspect Biol. 2020. PMID: 31501195 Free PMC article. Review.
-
Activated inositol 1,4,5-trisphosphate receptors are modified by homogeneous Lys-48- and Lys-63-linked ubiquitin chains, but only Lys-48-linked chains are required for degradation.J Biol Chem. 2011 Jan 14;286(2):1074-82. doi: 10.1074/jbc.M110.188383. Epub 2010 Nov 11. J Biol Chem. 2011. PMID: 21071436 Free PMC article.
-
The Targeting of Native Proteins to the Endoplasmic Reticulum-Associated Degradation (ERAD) Pathway: An Expanding Repertoire of Regulated Substrates.Biomolecules. 2021 Aug 11;11(8):1185. doi: 10.3390/biom11081185. Biomolecules. 2021. PMID: 34439852 Free PMC article. Review.
-
Transcriptome and Literature Mining Highlight the Differential Expression of ERLIN1 in Immune Cells during Sepsis.Biology (Basel). 2021 Aug 5;10(8):755. doi: 10.3390/biology10080755. Biology (Basel). 2021. PMID: 34439987 Free PMC article.
-
A VCP inhibitor substrate trapping approach (VISTA) enables proteomic profiling of endogenous ERAD substrates.Mol Biol Cell. 2018 May 1;29(9):1021-1030. doi: 10.1091/mbc.E17-08-0514. Epub 2018 Mar 22. Mol Biol Cell. 2018. PMID: 29514927 Free PMC article.
References
-
- Taylor CW, da Fonseca PCA, Morris EP. IP3 receptors: the search for structure. Trends Biochem. Sci. 2004;29:210–219. - PubMed
-
- Mikoshiba K. IP3 receptor/Ca2+ channel: from discovery to new signaling concepts. J. Neurochem. 2007;102:1426–1446. - PubMed
-
- Chan J, Whitten AE, Jeffries CM, Bosanac I, Mal TK, Ito J, Porumb H, Michikawa T, Mikoshiba K, Trewhella J, Ikura M. Ligand-induced conformational changes via flexible linkers in the amino-terminal region of the inositol 1,4,5-trisphosphate receptor. J. Mol. Biol. 2007;373:1269–1280. - PubMed
-
- Hamada K, Terauchi A, Mikoshiba K. Three-dimensional rearrangements within inositol 1,4,5-trisphosphate receptor by calcium. J. Biol. Chem. 2003;278:52881–52889. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
