

Evidence that a C1q/C1qR system regulates monocyte-derived dendritic cell differentiation at the interface of innate and acquired immunity
- PMID: 19710097
- PMCID: PMC2846191
- DOI: 10.1177/1753425909339815
Evidence that a C1q/C1qR system regulates monocyte-derived dendritic cell differentiation at the interface of innate and acquired immunity
Abstract
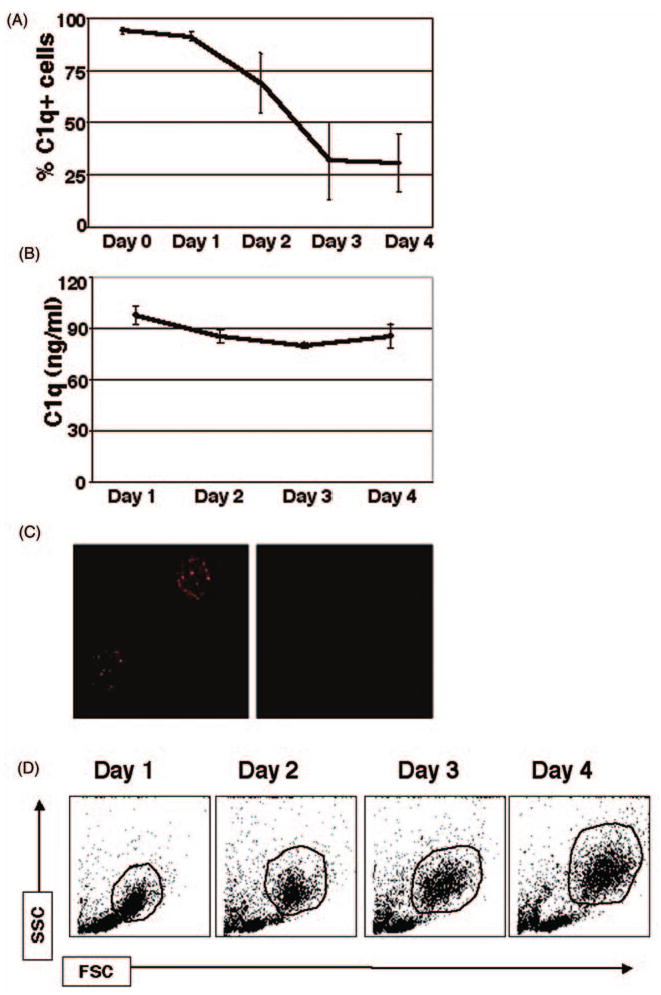
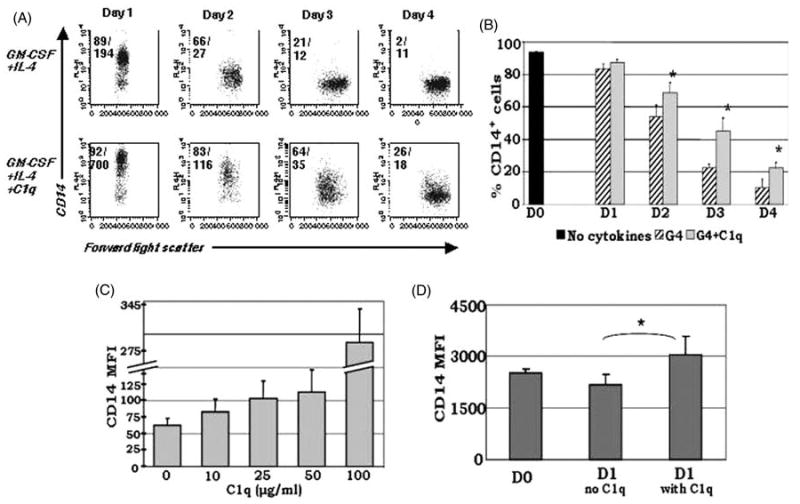
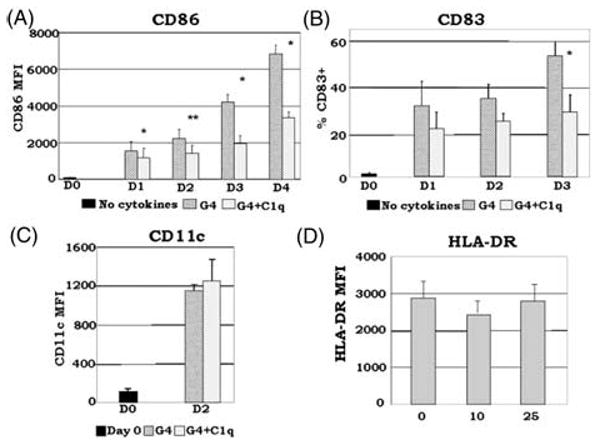
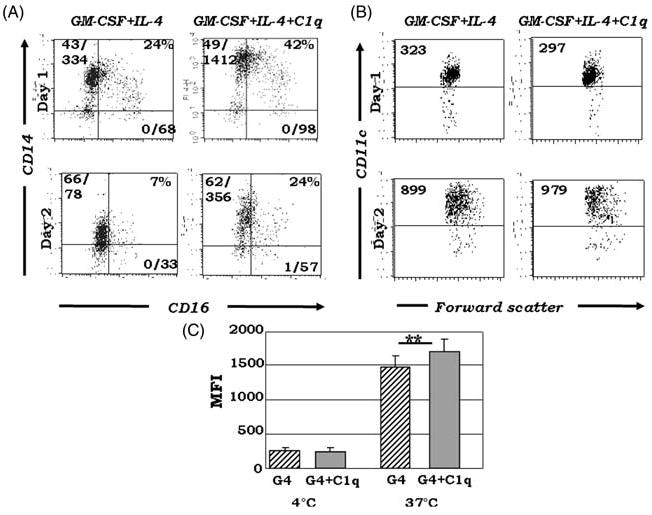
Growing evidence shows that C1q modulates the growth and function of cells committed to the monocyte-derived dendritic cell (DC) lineage. Because C1q regulates both innate and acquired immune responses, we postulated that C1q modulates the transition from monocytes to DCs, i.e. the interface between innate and acquired immunity. Human peripheral blood monocytes cultured with soluble C1q and DC growth factors (granulocyte-macrophage colony-stimulating factor + Interleukin-4) failed to down-regulate monocyte-associated (CD14, CD16) and up-regulate DC-associated (CD83, CD86) markers. Impaired DC differentiation was not due to apoptosis; further analysis revealed the development of CD14(hi)CD11c(hi)CD16 (+/-) cells that have previously been associated with both innate and acquired immunity. Monocyte-DC precursors expressed gC1qR, the receptor for globular heads of C1q, from the outset, while cC1qR, the receptor for the collagen tails of C1q, was expressed at low levels. Notably, the binding pattern of monoclonal antibodies specific to the globular heads of C1q indicated that C1q is bound to monocytes via globular heads, presumably through gC1qR. Moreover, gC1qR levels decreased, while cC1qR levels were dramatically amplified as monocytes differentiated into immature DC. Thus, specific C1q/C1q receptor (R) interactions may control the transition from the monocyte state (innate immunity) toward the professional antigen-presenting cell state (adaptive immunity).
Figures


Similar articles
-
SLE: Novel Postulates for Therapeutic Options.Front Immunol. 2020 Oct 7;11:583853. doi: 10.3389/fimmu.2020.583853. eCollection 2020. Front Immunol. 2020. PMID: 33117397 Free PMC article. Review.
-
Chemotaxis of human monocyte-derived dendritic cells to complement component C1q is mediated by the receptors gC1qR and cC1qR.Mol Immunol. 2006 Mar;43(9):1402-7. doi: 10.1016/j.molimm.2005.07.030. Epub 2005 Sep 2. Mol Immunol. 2006. PMID: 16140380
-
Maturation-dependent expression of C1q binding proteins on the cell surface of human monocyte-derived dendritic cells.Int Immunopharmacol. 2003 Jan;3(1):39-51. doi: 10.1016/s1567-5769(02)00211-4. Int Immunopharmacol. 2003. PMID: 12538033
-
DC-SIGN, C1q, and gC1qR form a trimolecular receptor complex on the surface of monocyte-derived immature dendritic cells.Blood. 2012 Aug 9;120(6):1228-36. doi: 10.1182/blood-2011-07-369728. Epub 2012 Jun 13. Blood. 2012. PMID: 22700724 Free PMC article.
-
Methods integrating innate and adaptive immune responses in human in vitro immunization assays.Front Immunol. 2025 May 21;16:1584852. doi: 10.3389/fimmu.2025.1584852. eCollection 2025. Front Immunol. 2025. PMID: 40469296 Free PMC article. Review.
Cited by
-
Role of the exogenous HCV core protein in the interaction of human hepatocyte proliferation and macrophage sub-populations.PLoS One. 2014 Sep 29;9(9):e108278. doi: 10.1371/journal.pone.0108278. eCollection 2014. PLoS One. 2014. PMID: 25265479 Free PMC article.
-
SLE: Novel Postulates for Therapeutic Options.Front Immunol. 2020 Oct 7;11:583853. doi: 10.3389/fimmu.2020.583853. eCollection 2020. Front Immunol. 2020. PMID: 33117397 Free PMC article. Review.
-
C1q Modulates the Response to TLR7 Stimulation by Pristane-Primed Macrophages: Implications for Pristane-Induced Lupus.J Immunol. 2016 Feb 15;196(4):1488-94. doi: 10.4049/jimmunol.1401009. Epub 2016 Jan 15. J Immunol. 2016. PMID: 26773156 Free PMC article.
-
The immunopathology of liver granulomas in primary biliary cirrhosis.J Autoimmun. 2012 Sep;39(3):216-21. doi: 10.1016/j.jaut.2012.05.022. Epub 2012 Jun 23. J Autoimmun. 2012. PMID: 22727562 Free PMC article.
-
Is the A-Chain the Engine That Drives the Diversity of C1q Functions? Revisiting Its Unique Structure.Front Immunol. 2018 Feb 5;9:162. doi: 10.3389/fimmu.2018.00162. eCollection 2018. Front Immunol. 2018. PMID: 29459870 Free PMC article.
References
-
- Steinman RM, Banchereau J. Taking dendritic cells into medicine. Nature. 2007;449:419–426. - PubMed
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
-
- Reid SD, Penna G, Adorini L. The control of T cell responses by dendritic cell subsets. Curr Opin Immunol. 2000;12:114–121. - PubMed
-
- Santiago-Schwarz F, Anand P, Liu S, Carsons SE. Dendritic cells (DCs) in rheumatoid arthritis (RA): progenitor cells and soluble factors contained in RA synovial fluid yield a subset of myeloid DCs that preferentially activate Th1 inflammatory-type responses. J Immunol. 2001;167:1758–1768. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
