

Ribose 2'-O methylation of the vesicular stomatitis virus mRNA cap precedes and facilitates subsequent guanine-N-7 methylation by the large polymerase protein
- PMID: 19710136
- PMCID: PMC2772757
- DOI: 10.1128/JVI.01426-09
Ribose 2'-O methylation of the vesicular stomatitis virus mRNA cap precedes and facilitates subsequent guanine-N-7 methylation by the large polymerase protein
Abstract
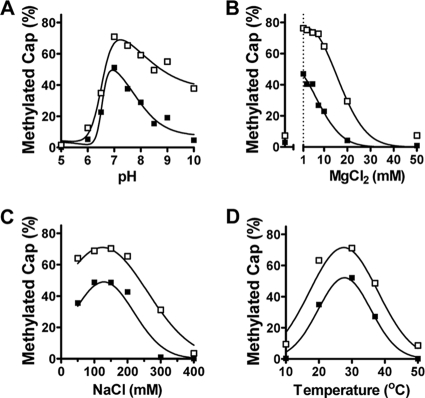
During conventional mRNA cap formation, two separate methyltransferases sequentially modify the cap structure, first at the guanine-N-7 (G-N-7) position and subsequently at the ribose 2'-O position. For vesicular stomatitis virus (VSV), a prototype of the nonsegmented negative-strand RNA viruses, the two methylase activities share a binding site for the methyl donor S-adenosyl-l-methionine and are inhibited by individual amino acid substitutions within the C-terminal domain of the large (L) polymerase protein. This led to the suggestion that a single methylase domain functions for both 2'-O and G-N-7 methylations. Here we report a trans-methylation assay that recapitulates both ribose 2'-O and G-N-7 modifications by using purified recombinant L and in vitro-synthesized RNA. Using this assay, we demonstrate that VSV L typically modifies the 2'-O position of the cap prior to the G-N-7 position and that G-N-7 methylation is diminished by pre-2'-O methylation of the substrate RNA. Amino acid substitutions in the C terminus of L that prevent all cap methylation in recombinant VSV (rVSV) partially retain the ability to G-N-7 methylate a pre-2'-O-methylated RNA, therefore uncoupling the effect of substitutions in the C terminus of the L protein on the two methylations. In addition, we show that the 2'-O and G-N-7 methylase activities act specifically on RNA substrates that contain the conserved elements of a VSV mRNA start at the 5' terminus. This study provides new mechanistic insights into the mRNA cap methylase activities of VSV L, demonstrates that 2'-O methylation precedes and facilitates subsequent G-N-7 methylation, and reveals an RNA sequence and length requirement for the two methylase activities. We propose a model of regulation of the activity of the C terminus of L protein in 2'-O and G-N-7 methylation of the cap structure.
Figures





Similar articles
-
Identification of aromatic amino acid residues in conserved region VI of the large polymerase of vesicular stomatitis virus is essential for both guanine-N-7 and ribose 2'-O methyltransferases.Virology. 2010 Dec 20;408(2):241-52. doi: 10.1016/j.virol.2010.09.017. Epub 2010 Oct 18. Virology. 2010. PMID: 20961592 Free PMC article.
-
A unique strategy for mRNA cap methylation used by vesicular stomatitis virus.Proc Natl Acad Sci U S A. 2006 May 30;103(22):8493-8. doi: 10.1073/pnas.0509821103. Epub 2006 May 18. Proc Natl Acad Sci U S A. 2006. PMID: 16709677 Free PMC article.
-
Vesicular stomatitis viruses resistant to the methylase inhibitor sinefungin upregulate RNA synthesis and reveal mutations that affect mRNA cap methylation.J Virol. 2007 Apr;81(8):4104-15. doi: 10.1128/JVI.02681-06. Epub 2007 Feb 14. J Virol. 2007. PMID: 17301155 Free PMC article.
-
An unconventional pathway of mRNA cap formation by vesiculoviruses.Virus Res. 2011 Dec;162(1-2):100-9. doi: 10.1016/j.virusres.2011.09.012. Epub 2011 Sep 16. Virus Res. 2011. PMID: 21945214 Free PMC article. Review.
-
Flavivirus methyltransferase: a novel antiviral target.Antiviral Res. 2008 Oct;80(1):1-10. doi: 10.1016/j.antiviral.2008.05.003. Epub 2008 Jun 5. Antiviral Res. 2008. PMID: 18571739 Free PMC article. Review.
Cited by
-
Sequence-function analysis of the Sendai virus L protein domain VI.Virology. 2010 Sep 30;405(2):370-82. doi: 10.1016/j.virol.2010.06.019. Epub 2010 Jul 6. Virology. 2010. PMID: 20609457 Free PMC article.
-
Molecular architecture of the vesicular stomatitis virus RNA polymerase.Proc Natl Acad Sci U S A. 2010 Nov 16;107(46):20075-80. doi: 10.1073/pnas.1013559107. Epub 2010 Nov 1. Proc Natl Acad Sci U S A. 2010. PMID: 21041632 Free PMC article.
-
Methylation of viral mRNA cap structures by PCIF1 attenuates the antiviral activity of interferon-β.Proc Natl Acad Sci U S A. 2021 Jul 20;118(29):e2025769118. doi: 10.1073/pnas.2025769118. Proc Natl Acad Sci U S A. 2021. PMID: 34266951 Free PMC article.
-
The L-VP35 and L-L interaction domains reside in the amino terminus of the Ebola virus L protein and are potential targets for antivirals.Virology. 2013 Jul 5;441(2):135-45. doi: 10.1016/j.virol.2013.03.013. Epub 2013 Apr 11. Virology. 2013. PMID: 23582637 Free PMC article.
-
Status of antiviral therapeutics against rabies virus and related emerging lyssaviruses.Curr Opin Virol. 2019 Apr;35:1-13. doi: 10.1016/j.coviro.2018.12.009. Epub 2019 Feb 10. Curr Opin Virol. 2019. PMID: 30753961 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
