

Bimolecular complementation of paramyxovirus fusion and hemagglutinin-neuraminidase proteins enhances fusion: implications for the mechanism of fusion triggering
- PMID: 19710150
- PMCID: PMC2772755
- DOI: 10.1128/JVI.01191-09
Bimolecular complementation of paramyxovirus fusion and hemagglutinin-neuraminidase proteins enhances fusion: implications for the mechanism of fusion triggering
Abstract
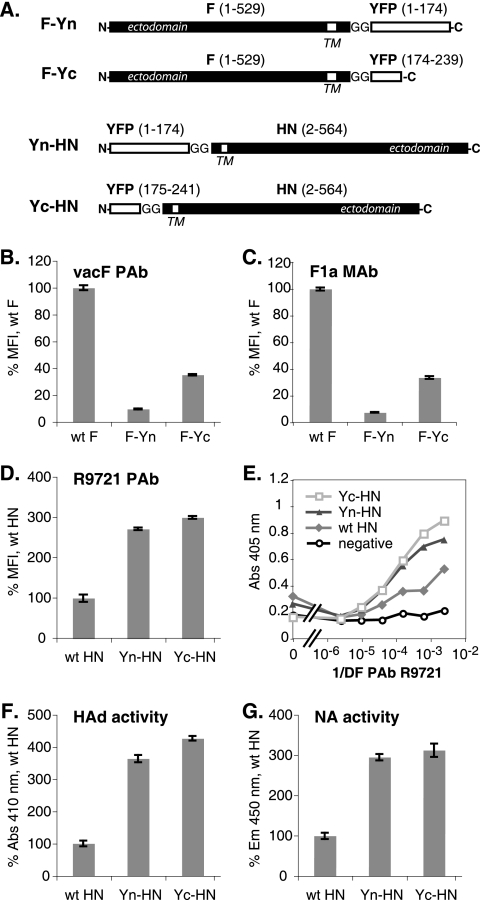
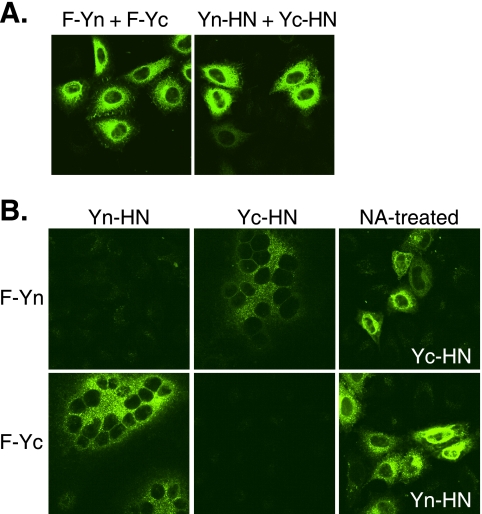
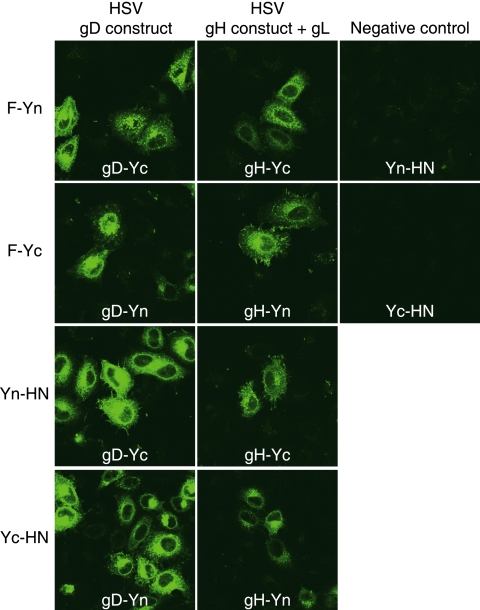
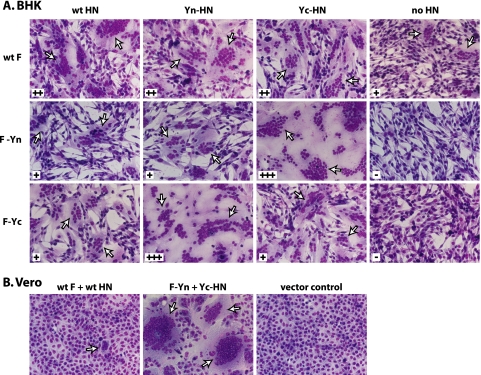
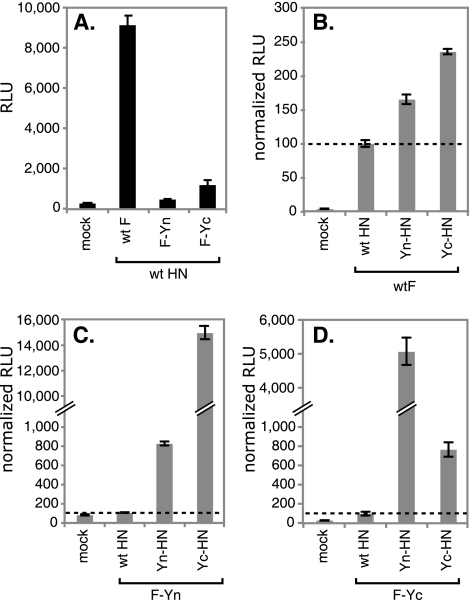
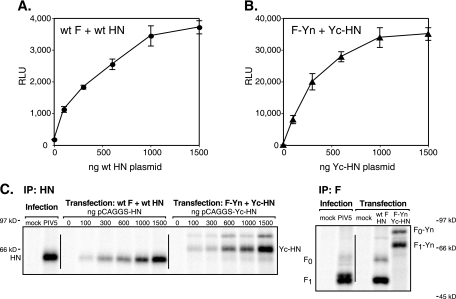
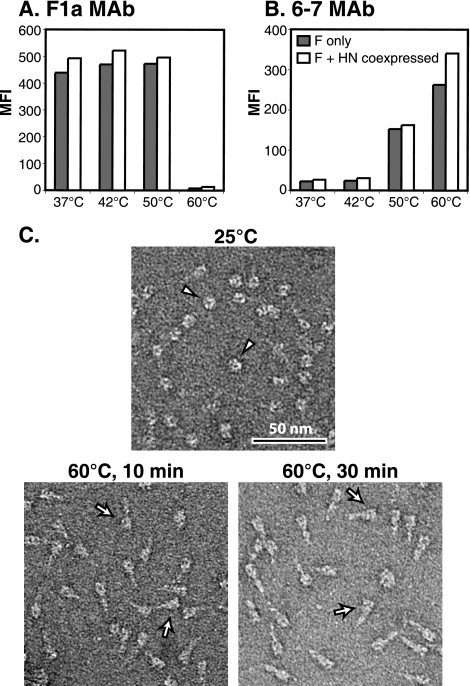
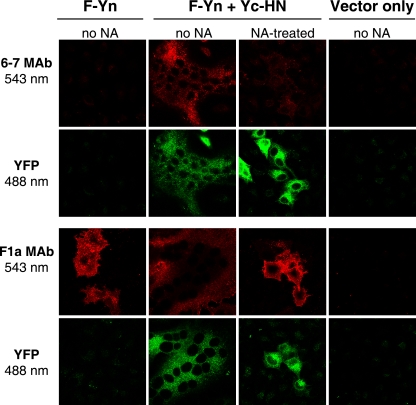
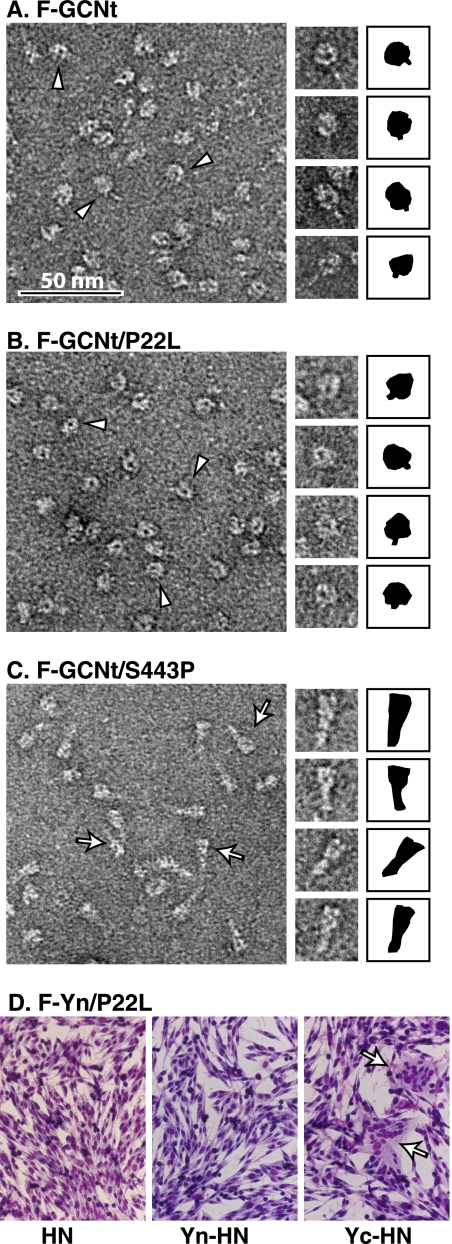
For paramyxoviruses, entry requires a receptor-binding protein (hemagglutinin-neuraminidase [HN], H, or G) and a fusion protein (F). Like other class I viral fusion proteins, F is expressed as a prefusion metastable protein that undergoes a refolding event to induce fusion. HN binding to its receptor triggers F refolding by an unknown mechanism. HN may serve as a clamp that stabilizes F in its prefusion state until HN binds the target cell (the "clamp model"). Alternatively, HN itself may undergo a conformational change after receptor binding that destabilizes F and causes F to trigger (the "provocateur model"). To examine F-HN interactions by bimolecular fluorescence complementation (BiFC), the cytoplasmic tails of parainfluenza virus 5 (PIV5) F and HN were fused to complementary fragments of yellow fluorescent protein (YFP). Coexpression of the BiFC constructs resulted in fluorescence; however, coexpression with unrelated BiFC constructs also produced fluorescence. The affinity of the two halves of YFP presumably superseded the F-HN interaction. Unexpectedly, coexpression of the BiFC F and HN constructs greatly enhanced fusion in multiple cell types. We hypothesize that the increase in fusion occurs because the BiFC tags bring F and HN together more frequently than occurs in a wild-type (wt) scenario. This implies that normally much of wt F is not associated with wt HN, in conflict with the clamp model for activation. Correspondingly, we show that wt PIV5 fusion occurs in an HN concentration-dependent manner. Also inconsistent with the clamp model are the findings that BiFC F does not adopt a postfusion conformation when expressed in the absence of HN and that HN coexpression does not provide resistance to the heat-induced triggering of F. In support of a provocateur model of F activation, we demonstrate by analysis of the morphology of soluble F trimers that the hyperfusogenic mutation S443P has a destabilizing effect on F.
Figures
References
-
- Aguilar, H. C., Z. A. Ataman, V. Aspericueta, A. Q. Fang, M. Stroud, O. A. Negrete, R. A. Kammerer, and B. Lee. 2009. A novel receptor-induced activation site in the Nipah virus attachment glycoprotein (G) involved in triggering the fusion glycoprotein (F). J. Biol. Chem. 284:1628-1635. - PMC - PubMed
-
- Aguilar, H. C., K. A. Matreyek, C. M. Filone, S. T. Hashimi, E. L. Levroney, O. A. Negrete, A. Betrolotti-Ciarlet, D. Y. Choi, I. McHardy, J. A. Fulcher, S. V. Su, M. C. Wolf, L. Kohatsu, L. G. Baum, and B. Lee. 2006. N-glycans on Nipah virus fusion protein protect against neutralization but reduce membrane fusion and viral entry. J. Virol. 80:4878-4889. - PMC - PubMed
-
- Bishop, K. A., A. C. Hickey, D. Khetawat, J. R. Patch, K. N. Bossart, Z. Zhu, L. F. Wang, D. S. Dimitrov, and C. C. Broder. 2008. Residues in the stalk domain of the Hendra virus G glycoprotein modulate conformational changes associated with receptor binding. J. Virol. 82:11398-11409. - PMC - PubMed
-
- Bishop, K. A., T. S. Stantchev, A. C. Hickey, D. Khetawat, K. N. Bossart, V. Krasnoperov, P. Gill, Y. R. Feng, L. Wang, B. T. Eaton, L. F. Wang, and C. C. Broder. 2007. Identification of Hendra virus G glycoprotein residues that are critical for receptor binding. J. Virol. 81:5893-5901. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
