

Probing PML body function in ALT cells reveals spatiotemporal requirements for telomere recombination
- PMID: 19717459
- PMCID: PMC2747187
- DOI: 10.1073/pnas.0907689106
Probing PML body function in ALT cells reveals spatiotemporal requirements for telomere recombination
Abstract
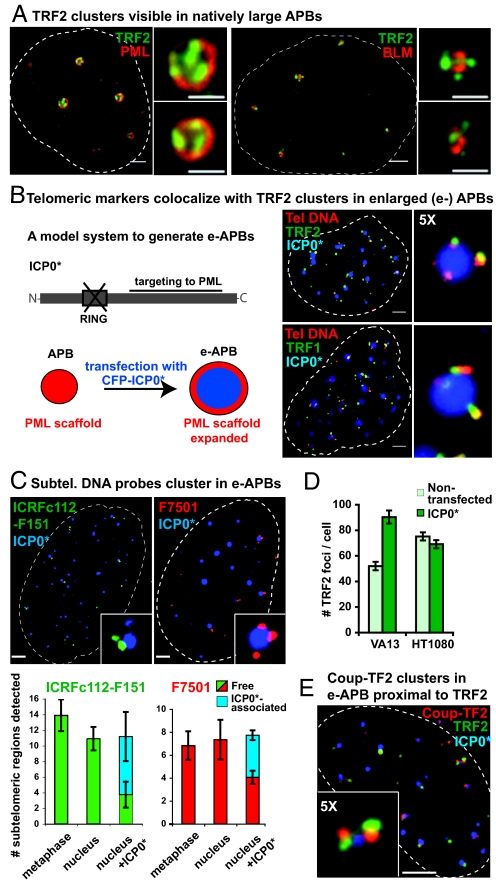
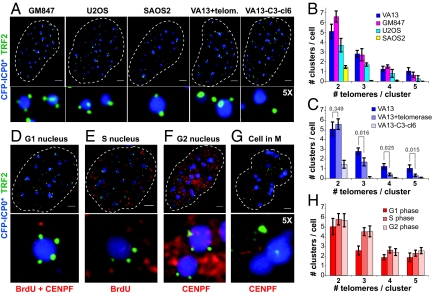
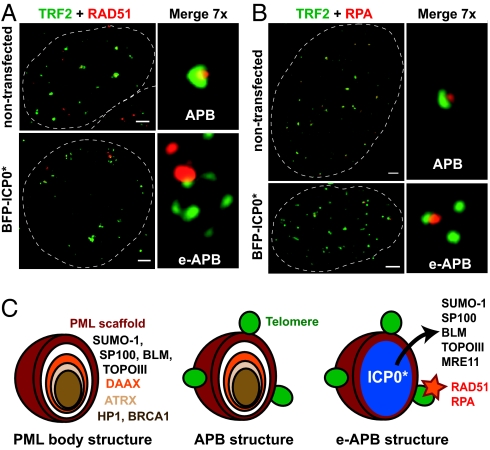
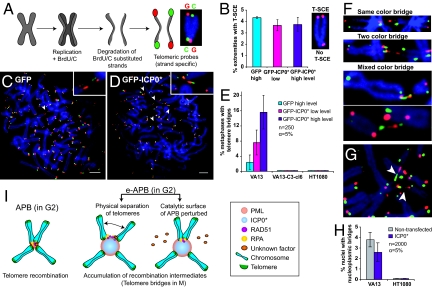
Promyelocytic leukemia (PML) bodies (also called ND10) are dynamic nuclear structures implicated in a wide variety of cellular processes. ALT-associated PML bodies (APBs) are specialized PML bodies found exclusively in telomerase-negative tumors in which telomeres are maintained by recombination-based alternative (ALT) mechanisms. Although it has been suggested that APBs are directly implicated in telomere metabolism of ALT cells, their precise role and structure have remained elusive. Here we show that PML bodies in ALT cells associate with chromosome ends forming small, spatially well-defined clusters, containing on average 2-5 telomeres. Using an innovative approach that gently enlarges PML bodies in living cells while retaining their overall organization, we show that this physical enlargement of APBs spatially resolves the single telomeres in the cluster, but does not perturb the potential of the APB to recruit chromosome extremities. We show that telomere clustering in PML bodies is cell-cycle regulated and that unique telomeres within a cluster associate with recombination proteins. Enlargement of APBs induced the accumulation of telomere-telomere recombination intermediates visible on metaphase spreads and connecting heterologous chromosomes. The strand composition of these recombination intermediates indicated that this recombination is constrained to a narrow time window in the cell cycle following replication. These data provide strong evidence that PML bodies are not only a marker for ALT cells but play a direct role in telomere recombination, both by bringing together chromosome ends and by promoting telomere-telomere interactions between heterologous chromosomes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bernardi R, Pandolfi PP. Structure, dynamics, and functions of promyelocytic leukaemia nuclear bodies. Nat Rev Mol Cell Biol. 2007;8:1006–1016. - PubMed
-
- Yeager TR, et al. Telomerase-negative immortalized human cells contain a novel type of promyelocytic leukemia (PML) body. Cancer Res. 1999;59:4175–4179. - PubMed
-
- Jiang WQ, Zhong ZH, Henson JD, Reddel RR. Identification of candidate alternative lengthening of telomeres genes by methionine restriction and RNA interference. Oncogene. 2007;26:4635–4647. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
