

Structural basis of AdoMet-dependent aminocarboxypropyl transfer reaction catalyzed by tRNA-wybutosine synthesizing enzyme, TYW2
- PMID: 19717466
- PMCID: PMC2747168
- DOI: 10.1073/pnas.0905270106
Structural basis of AdoMet-dependent aminocarboxypropyl transfer reaction catalyzed by tRNA-wybutosine synthesizing enzyme, TYW2
Abstract
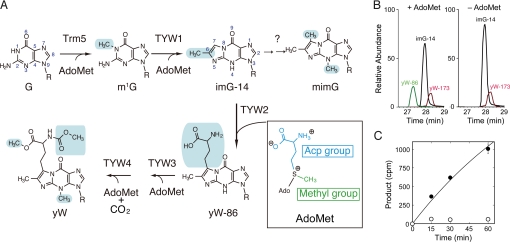
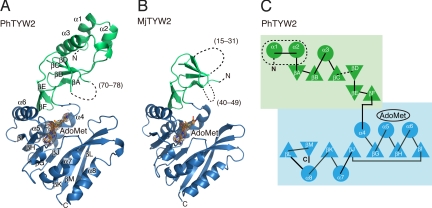
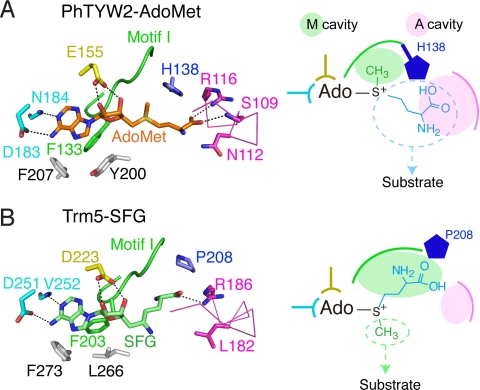
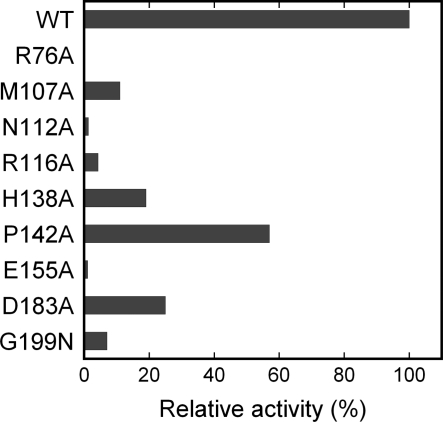
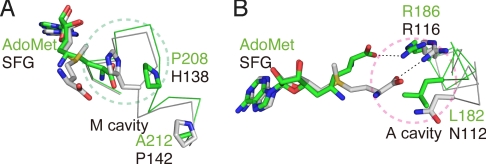
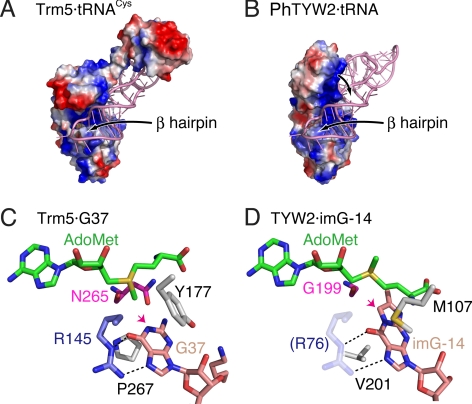
S-adenosylmethionine (AdoMet) is a methyl donor used by a wide variety of methyltransferases, and it is also used as the source of an alpha-amino-alpha-carboxypropyl ("acp") group by several enzymes. tRNA-yW synthesizing enzyme-2 (TYW2) is involved in the biogenesis of a hypermodified nucleotide, wybutosine (yW), and it catalyzes the transfer of the "acp" group from AdoMet to the C7 position of the imG-14 base, a yW precursor. This modified nucleoside yW is exclusively located at position 37 of eukaryotic tRNA(Phe), and it ensures the anticodon-codon pairing on the ribosomal decoding site. Although this "acp" group has a significant role in preventing decoding frame shifts, the mechanism of the "acp" group transfer by TYW2 remains unresolved. Here we report the crystal structures and functional analyses of two archaeal homologs of TYW2 from Pyrococcus horikoshii and Methanococcus jannaschii. The in vitro mass spectrometric and radioisotope-labeling analyses confirmed that these archaeal TYW2 homologues have the same activity as yeast TYW2. The crystal structures verified that the archaeal TYW2 contains a canonical class-I methyltransferase (MTase) fold. However, their AdoMet-bound structures revealed distinctive AdoMet-binding modes, in which the "acp" group, instead of the methyl group, of AdoMet is directed to the substrate binding pocket. Our findings, which were confirmed by extensive mutagenesis studies, explain why TYW2 transfers the "acp" group, and not the methyl group, from AdoMet to the nucleobase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structure-function analysis of human TYW2 enzyme required for the biosynthesis of a highly modified Wybutosine (yW) base in phenylalanine-tRNA.PLoS One. 2012;7(6):e39297. doi: 10.1371/journal.pone.0039297. Epub 2012 Jun 28. PLoS One. 2012. PMID: 22761755 Free PMC article.
-
Biosynthesis of wyosine derivatives in tRNA(Phe) of Archaea: role of a remarkable bifunctional tRNA(Phe):m1G/imG2 methyltransferase.RNA. 2014 Jun;20(6):747-53. doi: 10.1261/rna.043315.113. RNA. 2014. PMID: 24837075 Free PMC article. Review.
-
Structural basis of tRNA modification with CO2 fixation and methylation by wybutosine synthesizing enzyme TYW4.Nucleic Acids Res. 2009 May;37(9):2910-25. doi: 10.1093/nar/gkp158. Epub 2009 Mar 14. Nucleic Acids Res. 2009. PMID: 19287006 Free PMC article.
-
Biosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNA.EMBO J. 2006 May 17;25(10):2142-54. doi: 10.1038/sj.emboj.7601105. Epub 2006 Apr 27. EMBO J. 2006. PMID: 16642040 Free PMC article.
-
Wybutosine biosynthesis: structural and mechanistic overview.RNA Biol. 2014;11(12):1508-18. doi: 10.4161/15476286.2014.992271. RNA Biol. 2014. PMID: 25629788 Free PMC article. Review.
Cited by
-
Structural and functional characterization of the TYW3/Taw3 class of SAM-dependent methyltransferases.RNA. 2017 Mar;23(3):346-354. doi: 10.1261/rna.057943.116. Epub 2016 Dec 8. RNA. 2017. PMID: 27932585 Free PMC article.
-
Biosynthesis of wyosine derivatives in tRNA: an ancient and highly diverse pathway in Archaea.Mol Biol Evol. 2010 Sep;27(9):2062-77. doi: 10.1093/molbev/msq096. Epub 2010 Apr 9. Mol Biol Evol. 2010. PMID: 20382657 Free PMC article.
-
Structure-function analysis of human TYW2 enzyme required for the biosynthesis of a highly modified Wybutosine (yW) base in phenylalanine-tRNA.PLoS One. 2012;7(6):e39297. doi: 10.1371/journal.pone.0039297. Epub 2012 Jun 28. PLoS One. 2012. PMID: 22761755 Free PMC article.
-
Biosynthesis of wyosine derivatives in tRNA(Phe) of Archaea: role of a remarkable bifunctional tRNA(Phe):m1G/imG2 methyltransferase.RNA. 2014 Jun;20(6):747-53. doi: 10.1261/rna.043315.113. RNA. 2014. PMID: 24837075 Free PMC article. Review.
-
Radical SAM enzymes involved in the biosynthesis of purine-based natural products.Biochim Biophys Acta. 2012 Nov;1824(11):1245-53. doi: 10.1016/j.bbapap.2012.07.014. Epub 2012 Aug 3. Biochim Biophys Acta. 2012. PMID: 22902275 Free PMC article. Review.
References
-
- Ishitani R, Yokoyama S, Nureki O. Structure, dynamics, and function of RNA modification enzymes. Curr Opin Struct Biol. 2008;18:330–339. - PubMed
-
- Blobstein SH, Grunberger D, Weinstein IB, Nakanishi K. Isolation and structure determination of the fluorescent base from bovine liver phenylalanine transfer ribonucleic acid. Biochemistry. 1973;12:188–193. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
