

beta6 Integrin subunit deficiency alleviates lung injury in a mouse model of bronchopulmonary dysplasia
- PMID: 19717813
- PMCID: PMC2911573
- DOI: 10.1165/rcmb.2008-0480OC
beta6 Integrin subunit deficiency alleviates lung injury in a mouse model of bronchopulmonary dysplasia
Abstract
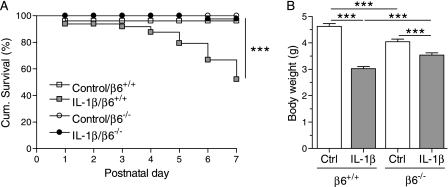
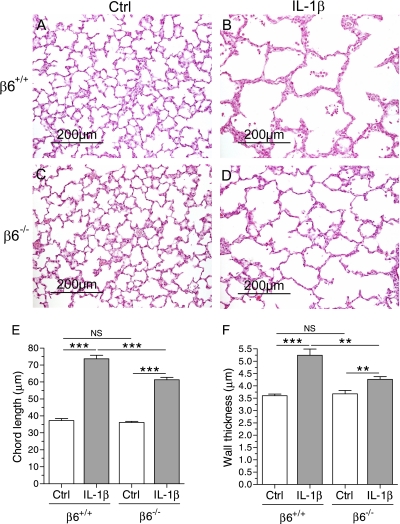
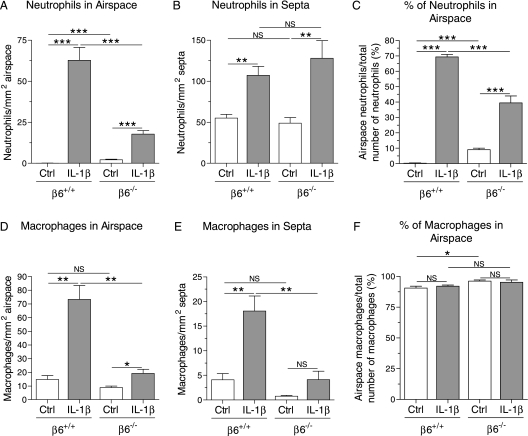
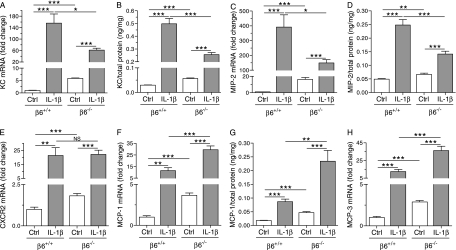
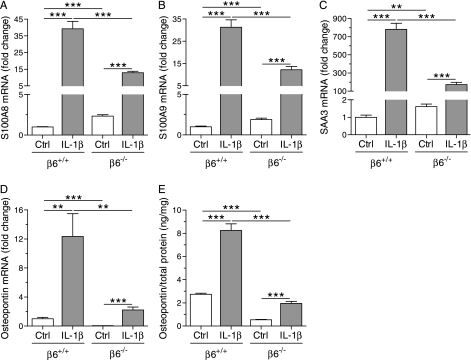
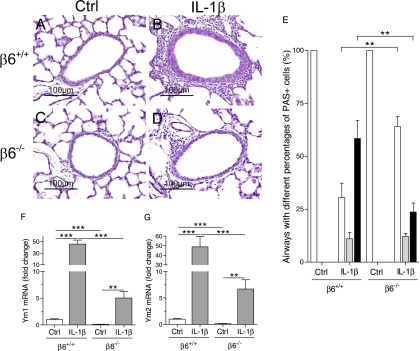
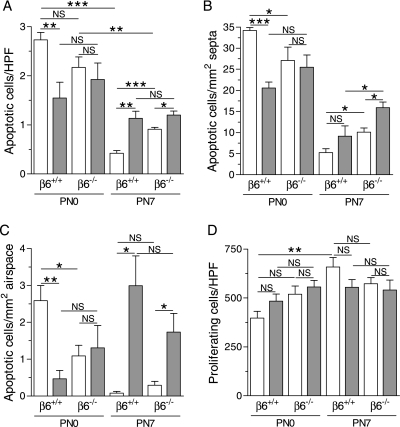
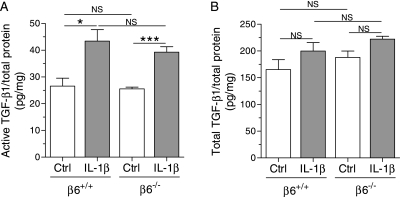
Pulmonary inflammation is associated with the development of bronchopulmonary dysplasia in premature infants. We have previously shown that perinatal pulmonary expression of human IL-1beta is sufficient to cause a lung disease similar to bronchopulmonary dysplasia, characterized by inflammation, impaired alveolarization, poor postnatal growth, and increased mortality in infant mice. The alphavbeta6 integrin plays a critical role in regulating inflammation in the adult lung. To study the role of the beta6 integrin subunit in neonatal inflammatory lung disease, we compared the pulmonary development in IL-1beta-expressing infant mice with wild-type or null beta6 integrin loci. Absence of the beta6 integrin subunit decreased the mortality and improved the postnatal growth of IL-1beta-expressing pups. The disrupted alveolar development of IL-1beta-expressing mice was improved by beta6 integrin deficiency. IL-1beta-expressing beta6(-/-) pups had shorter alveolar chord length and thinner alveolar walls than IL-1beta-expressing beta6(+/+) pups. In addition, the absence of the beta6 integrin subunit reduced IL-1beta-induced neutrophil and macrophage infiltration into the alveolar spaces. beta6 integrin subunit deficiency suppressed inflammation and goblet cell hyperplasia in the airways and alleviated airway remodeling in IL-1beta-expressing mice. The expression of the chemoattractant proteins, keratinocyte-derived chemokine, macrophage-inflammatory protein-2, calgranulin A, and calgranulin B, of osteopontin, and of the chitinase-like lectins, Ym1 and Ym2, was lower in IL-1beta-expressing beta6(-/-) than in IL-1beta-expressing beta6(+/+) mice. We conclude that absence of the beta6 integrin subunit protects the infant murine lung against IL-1beta-induced inflammation and injury.
Figures
Similar articles
-
Mechanisms of inflammatory lung injury in the neonate: lessons from a transgenic mouse model of bronchopulmonary dysplasia.Semin Perinatol. 2010 Jun;34(3):211-21. doi: 10.1053/j.semperi.2010.02.006. Semin Perinatol. 2010. PMID: 20494738 Review.
-
Role of CXC chemokine receptor-2 in a murine model of bronchopulmonary dysplasia.Am J Respir Cell Mol Biol. 2012 Dec;47(6):746-58. doi: 10.1165/rcmb.2011-0394OC. Epub 2012 Aug 3. Am J Respir Cell Mol Biol. 2012. PMID: 22865624
-
Maternal IL-1beta production prevents lung injury in a mouse model of bronchopulmonary dysplasia.Am J Respir Cell Mol Biol. 2010 Feb;42(2):149-60. doi: 10.1165/rcmb.2008-0287OC. Epub 2009 May 1. Am J Respir Cell Mol Biol. 2010. PMID: 19411613 Free PMC article.
-
Developmental stage is a major determinant of lung injury in a murine model of bronchopulmonary dysplasia.Pediatr Res. 2011 Apr;69(4):312-8. doi: 10.1203/PDR.0b013e31820bcb2a. Pediatr Res. 2011. PMID: 21178818
-
Halogen exposure injury in the developing lung.Ann N Y Acad Sci. 2020 Nov;1480(1):30-43. doi: 10.1111/nyas.14445. Epub 2020 Aug 1. Ann N Y Acad Sci. 2020. PMID: 32738176 Free PMC article. Review.
Cited by
-
Understanding the Impact of Infection, Inflammation, and Their Persistence in the Pathogenesis of Bronchopulmonary Dysplasia.Front Med (Lausanne). 2015 Dec 21;2:90. doi: 10.3389/fmed.2015.00090. eCollection 2015. Front Med (Lausanne). 2015. PMID: 26734611 Free PMC article. Review.
-
Impaired Myofibroblast Proliferation is a Central Feature of Pathologic Post-Natal Alveolar Simplification.bioRxiv [Preprint]. 2024 Sep 16:2023.12.21.572766. doi: 10.1101/2023.12.21.572766. bioRxiv. 2024. Update in: Elife. 2024 Dec 11;13:RP94425. doi: 10.7554/eLife.94425. PMID: 38187712 Free PMC article. Updated. Preprint.
-
Titanium oxide nanoparticle instillation induces inflammation and inhibits lung development in mice.Am J Physiol Lung Cell Mol Physiol. 2013 Feb 1;304(3):L152-61. doi: 10.1152/ajplung.00013.2012. Epub 2012 Dec 7. Am J Physiol Lung Cell Mol Physiol. 2013. PMID: 23220372 Free PMC article.
-
Distinct gene loci control the host response to influenza H1N1 virus infection in a time-dependent manner.BMC Genomics. 2012 Aug 20;13:411. doi: 10.1186/1471-2164-13-411. BMC Genomics. 2012. PMID: 22905720 Free PMC article.
-
Do cell junction protein mutations cause an airway phenotype in mice or humans?Am J Respir Cell Mol Biol. 2011 Aug;45(2):202-20. doi: 10.1165/rcmb.2010-0498TR. Epub 2011 Feb 4. Am J Respir Cell Mol Biol. 2011. PMID: 21297078 Free PMC article. Review.
References
-
- Jobe AH, Bancalari E. Bronchopulmonary dysplasia. Am J Respir Crit Care Med 2001;163:1723–1729. - PubMed
-
- Lemons JA, Bauer CR, Oh W, Korones SB, Papile L-A, Stoll BJ, Verter J, Temprosa M, Wright LL, Ehrenkranz RA, et al. Very low birth weight outcomes of the National Institute of Child Health and Human Development Neonatal Research Network, January 1995 through December 1996. Pediatrics 2001;107:e1. - PubMed
-
- Coalson JJ. Pathology of new bronchopulmonary dysplasia. Semin Neonatol 2003;8:73–81. - PubMed
-
- Goldenberg RL, Hauth JC, Andrews WW. Intrauterine infection and preterm delivery. N Engl J Med 2000;342:1500–1507. - PubMed
-
- Watterberg KL, Demers LM, Scott SM, Murphy S. Chorioamnionitis and early lung inflammation in infants in whom bronchopulmonary dysplasia develops. Pediatrics 1996;97:210–215. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
