

Tissue regenerative delays and synthetic lethality in adult mice after combined deletion of Atr and Trp53
- PMID: 19718024
- PMCID: PMC2823374
- DOI: 10.1038/ng.441
Tissue regenerative delays and synthetic lethality in adult mice after combined deletion of Atr and Trp53
Abstract
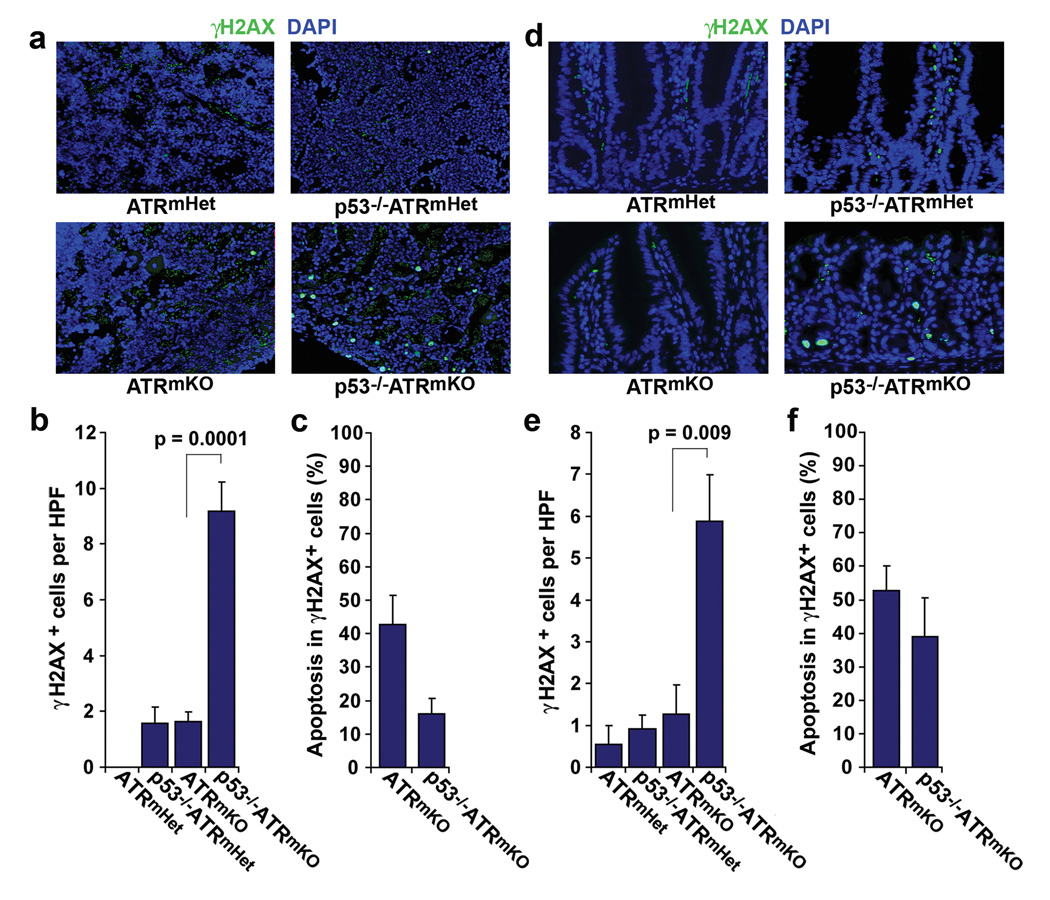
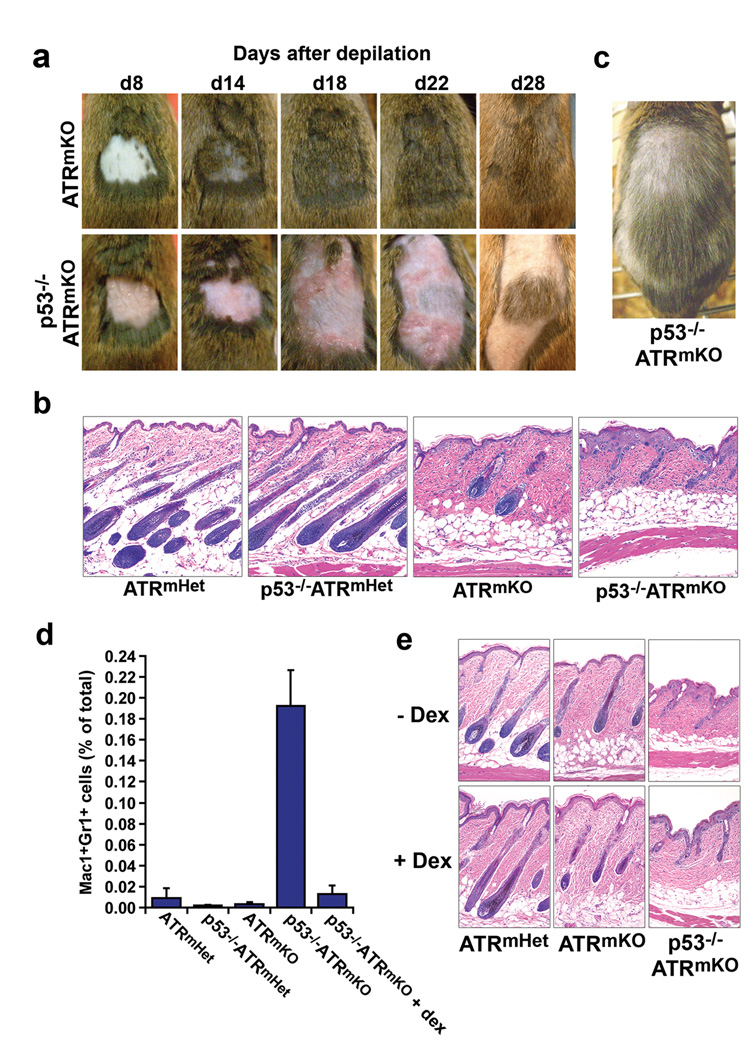
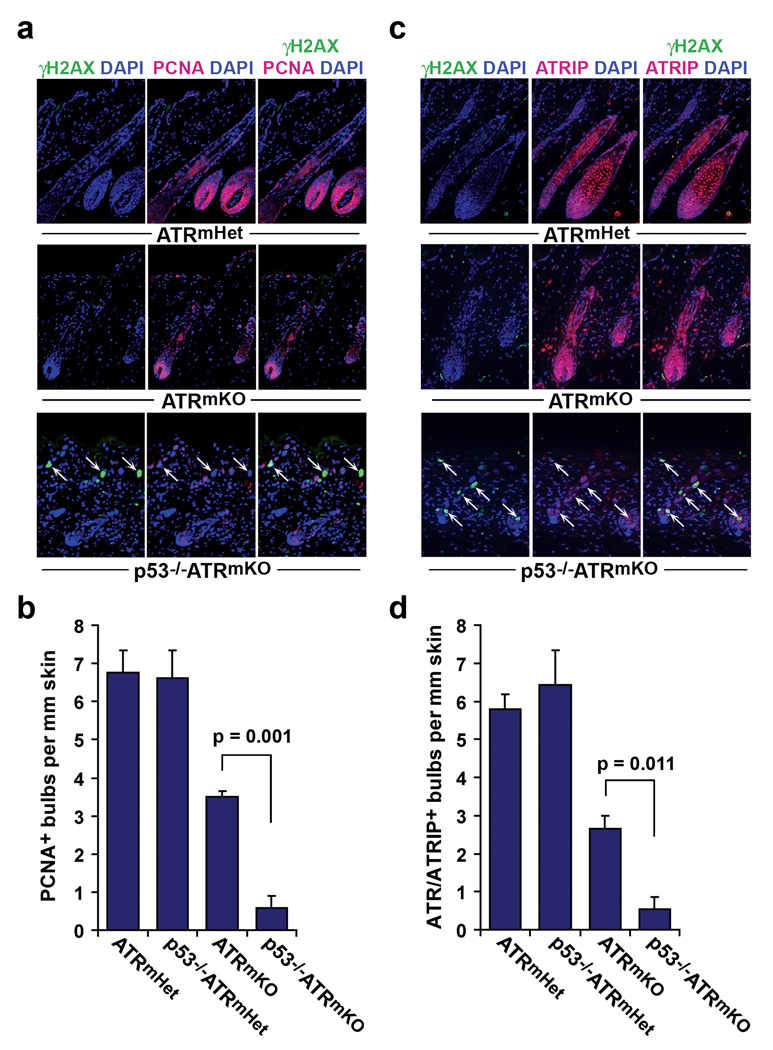
Trp53 loss of function has previously been shown to rescue tissue maintenance and developmental defects resulting from DNA damage or DNA-repair gene mutations. Here, we report that p53 deficiency severely exacerbates tissue degeneration caused by mosaic deletion of the essential genome maintenance regulator Atr. Combined loss of Atr and p53 (Trp53(-/-)Atr(mKO)) led to severe defects in hair follicle regeneration, localized inflammation (Mac1(+)Gr1(+) infiltrates), accelerated deterioration of the intestinal epithelium and synthetic lethality in adult mice. Tissue degeneration in Trp53(-/-)Atr(mKO) mice was characterized by the accumulation of cells maintaining high levels of DNA damage. Moreover, the elevated frequency of these damaged cells in both progenitor and downstream compartments in Trp53(-/-)Atr(mKO) skin coincided with delayed compensatory tissue renewal from residual ATR-expressing cells. Together, our results indicate that the combined loss of Atr and Trp53 in adult mice leads to the accumulation of highly damaged cells, which, consequently, impose a barrier to regeneration from undamaged progenitors.
Figures


References
-
- Wlodarski P, et al. Role of p53 in hematopoietic recovery after cytotoxic treatment. Blood. 1998;91:2998–3006. - PubMed
-
- Chin L, et al. p53 deficiency rescues the adverse effects of telomere loss and cooperates with telomere dysfunction to accelerate carcinogenesis. Cell. 1999;97:527–538. - PubMed
-
- Frank KM, et al. DNA ligase IV deficiency in mice leads to defective neurogenesis and embryonic lethality via the p53 pathway. Mol Cell. 2000;5:993–1002. - PubMed
-
- Gao Y, et al. Interplay of p53 and DNA-repair protein XRCC4 in tumorigenesis, genomic stability and development. Nature. 2000;404:897–900. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
