

The evolution of eyes and visually guided behaviour
- PMID: 19720648
- PMCID: PMC2781862
- DOI: 10.1098/rstb.2009.0083
The evolution of eyes and visually guided behaviour
Abstract
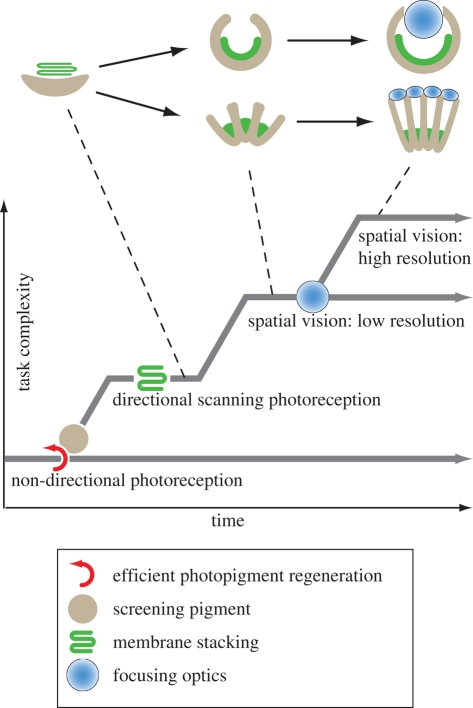
The morphology and molecular mechanisms of animal photoreceptor cells and eyes reveal a complex pattern of duplications and co-option of genetic modules, leading to a number of different light-sensitive systems that share many components, in which clear-cut homologies are rare. On the basis of molecular and morphological findings, I discuss the functional requirements for vision and how these have constrained the evolution of eyes. The fact that natural selection on eyes acts through the consequences of visually guided behaviour leads to a concept of task-punctuated evolution, where sensory systems evolve by a sequential acquisition of sensory tasks. I identify four key innovations that, one after the other, paved the way for the evolution of efficient eyes. These innovations are (i) efficient photopigments, (ii) directionality through screening pigment, (iii) photoreceptor membrane folding, and (iv) focusing optics. A corresponding evolutionary sequence is suggested, starting at non-directional monitoring of ambient luminance and leading to comparisons of luminances within a scene, first by a scanning mode and later by parallel spatial channels in imaging eyes.
Figures
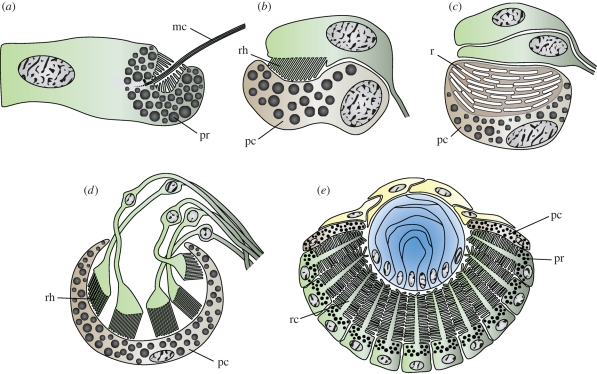
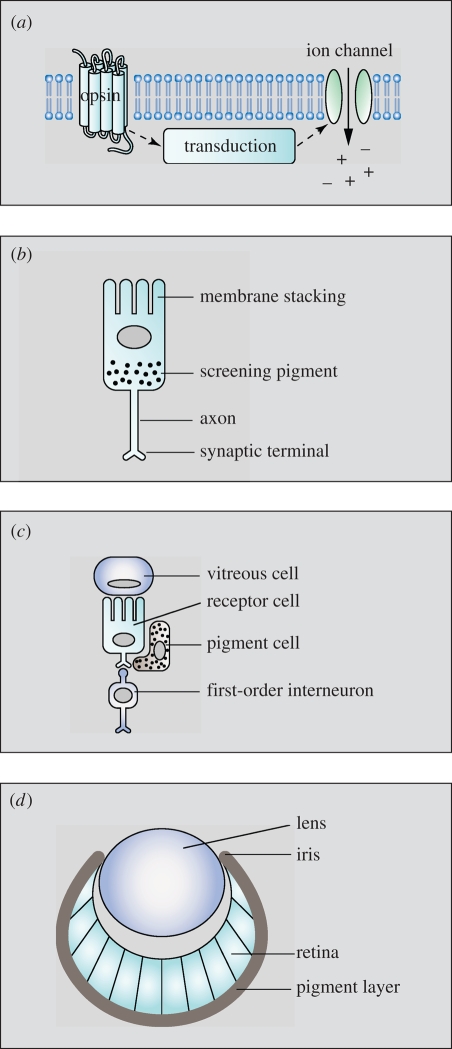
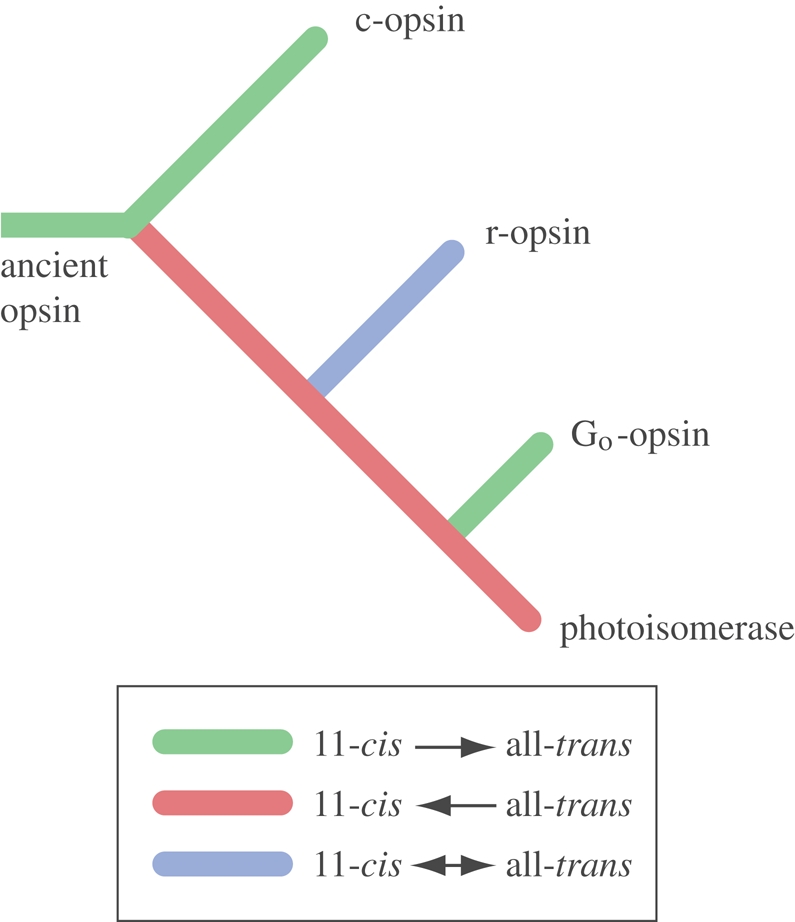
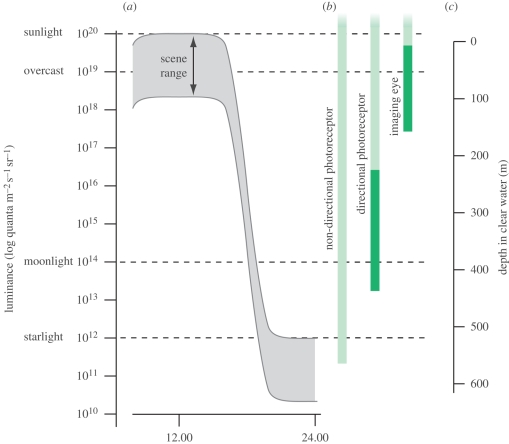
Similar articles
-
Eye evolution and its functional basis.Vis Neurosci. 2013 Mar;30(1-2):5-20. doi: 10.1017/S0952523813000035. Vis Neurosci. 2013. PMID: 23578808 Free PMC article.
-
Evolving eyes.Int J Dev Biol. 2004;48(8-9):701-5. doi: 10.1387/ijdb.041888rf. Int J Dev Biol. 2004. PMID: 15558462 Review.
-
Eye evolution: common use and independent recruitment of genetic components.Philos Trans R Soc Lond B Biol Sci. 2009 Oct 12;364(1531):2819-32. doi: 10.1098/rstb.2009.0079. Philos Trans R Soc Lond B Biol Sci. 2009. PMID: 19720647 Free PMC article. Review.
-
Amphioxus photoreceptors - insights into the evolution of vertebrate opsins, vision and circadian rhythmicity.Int J Dev Biol. 2017;61(10-11-12):665-681. doi: 10.1387/ijdb.170230zk. Int J Dev Biol. 2017. PMID: 29319115 Review.
-
Molecular Evidence for Convergence and Parallelism in Evolution of Complex Brains of Cephalopod Molluscs: Insights from Visual Systems.Integr Comp Biol. 2015 Dec;55(6):1070-83. doi: 10.1093/icb/icv049. Epub 2015 May 21. Integr Comp Biol. 2015. PMID: 26002349 Free PMC article.
Cited by
-
An empirical test of convergent evolution in rhodopsins.Mol Biol Evol. 2014 Jan;31(1):85-95. doi: 10.1093/molbev/mst171. Epub 2013 Sep 27. Mol Biol Evol. 2014. PMID: 24077848 Free PMC article.
-
Contrast sensitivity and the detection of moving patterns and features.Philos Trans R Soc Lond B Biol Sci. 2014 Jan 6;369(1636):20130043. doi: 10.1098/rstb.2013.0043. Print 2014. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 24395970 Free PMC article.
-
Eye evolution and its functional basis.Vis Neurosci. 2013 Mar;30(1-2):5-20. doi: 10.1017/S0952523813000035. Vis Neurosci. 2013. PMID: 23578808 Free PMC article.
-
dlx and sp6-9 Control optic cup regeneration in a prototypic eye.PLoS Genet. 2011 Aug;7(8):e1002226. doi: 10.1371/journal.pgen.1002226. Epub 2011 Aug 11. PLoS Genet. 2011. PMID: 21852957 Free PMC article.
-
High opsin diversity in a non-visual infaunal brittle star.BMC Genomics. 2014 Nov 28;15:1035. doi: 10.1186/1471-2164-15-1035. BMC Genomics. 2014. PMID: 25429842 Free PMC article.
References
-
- Arendt D.2003Evolution of eyes and photoreceptor cell types. Int. J. Dev. Biol. 47, 563–571 - PubMed
-
- Arendt D.2008The evolution of cell types in animals: emerging principles from molecular studies. Nat. Rev. Genet. 9, 868–882 (doi:10.1038/nrg2416) - DOI - PubMed
-
- Arendt D., Wittbrodt J.2001Reconstructing the eyes of Urbilateria. Phil. Trans. R. Soc. Lond. B 356, 1545–1563 (doi:10.1098/rstb.2001.0971) - DOI - PMC - PubMed
-
- Arendt D., Tessmar-Raible K., Snyman H., Dorresteijn A. W., Wittbrodt J.2004Ciliary photoreceptors with a vertebrate-type opsin in an invertebrate brain. Science 306, 869–871 (doi:10.1126/science.1099955) - DOI - PubMed
-
- Arendt D., Hausen H., Purschke G.2009The “division of labour” model of eye evolution. Phil. Trans. R. Soc. B 364, 2809–2817 (doi:10.1018/rstb.2009.0104) - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
