

The evolution of irradiance detection: melanopsin and the non-visual opsins
- PMID: 19720649
- PMCID: PMC2781857
- DOI: 10.1098/rstb.2009.0050
The evolution of irradiance detection: melanopsin and the non-visual opsins
Abstract
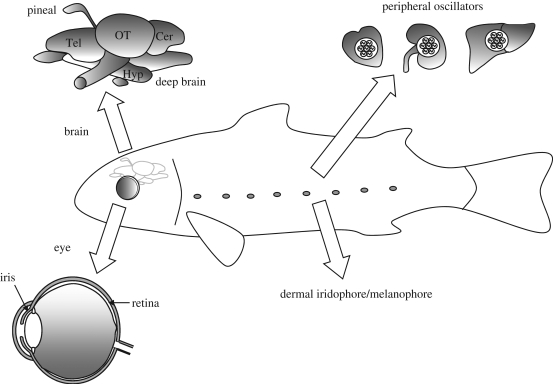
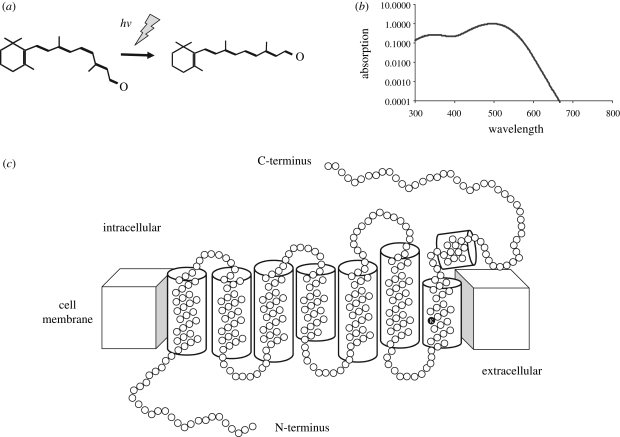
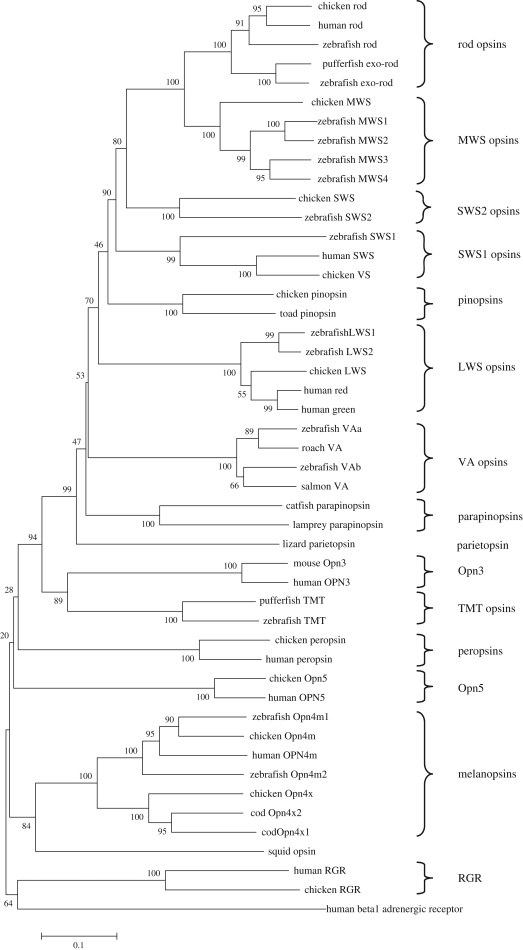
Circadian rhythms are endogenous 24 h cycles that persist in the absence of external time cues. These rhythms provide an internal representation of day length and optimize physiology and behaviour to the varying demands of the solar cycle. These clocks require daily adjustment to local time and the primary time cue (zeitgeber) used by most vertebrates is the daily change in the amount of environmental light (irradiance) at dawn and dusk, a process termed photoentrainment. Attempts to understand the photoreceptor mechanisms mediating non-image-forming responses to light, such as photoentrainment, have resulted in the discovery of a remarkable array of different photoreceptors and photopigment families, all of which appear to use a basic opsin/vitamin A-based photopigment biochemistry. In non-mammalian vertebrates, specialized photoreceptors are located within the pineal complex, deep brain and dermal melanophores. There is also strong evidence in fish and amphibians for the direct photic regulation of circadian clocks in multiple tissues. By contrast, mammals possess only ocular photoreceptors. However, in addition to the image-forming rods and cones of the retina, there exists a third photoreceptor system based on a subset of melanopsin-expressing photosensitive retinal ganglion cells (pRGCs). In this review, we discuss the range of vertebrate photoreceptors and their opsin photopigments, describe the melanopsin/pRGC system in some detail and then finally consider the molecular evolution and sensory ecology of these non-image-forming photoreceptor systems.
Figures
References
-
- Altimus C. M., Guler A. D., Villa K. L., McNeill D. S., Legates T. A., Hattar S.2008Rods–cones and melanopsin detect light and dark to modulate sleep independent of image formation. Proc. Natl Acad. Sci. USA 105, 19 998–20 003 (doi:10.1073/pnas.0808312105) - DOI - PMC - PubMed
-
- Arendt J.1998Melatonin and the pineal gland: influence on mammalian seasonal and circadian physiology. Rev. Reprod. 3, 13–22 (doi:10.1530/ror.0.0030013) - DOI - PubMed
-
- Arendt D.2003Evolution of eyes and photoreceptor cell types. Int. J. Dev. Biol. 47, 563–571 - PubMed
-
- Aschoff J.1984Circadian timing. Ann. N. Y. Acad. Sci. 423, 442–468 (doi:10.1111/j.1749-6632.1984.tb23452.x) - DOI - PubMed
-
- Barlow H. B.1957Purkinje shift and retinal noise. Nature 179, 255–256 (doi:10.1038/179255b0) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources