

Evolution of vertebrate rod and cone phototransduction genes
- PMID: 19720650
- PMCID: PMC2781860
- DOI: 10.1098/rstb.2009.0077
Evolution of vertebrate rod and cone phototransduction genes
Abstract
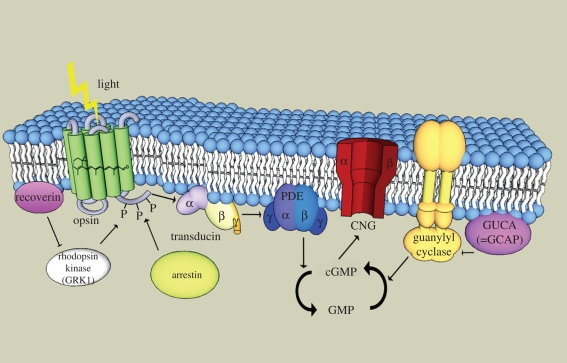
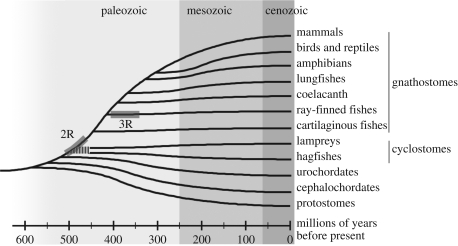
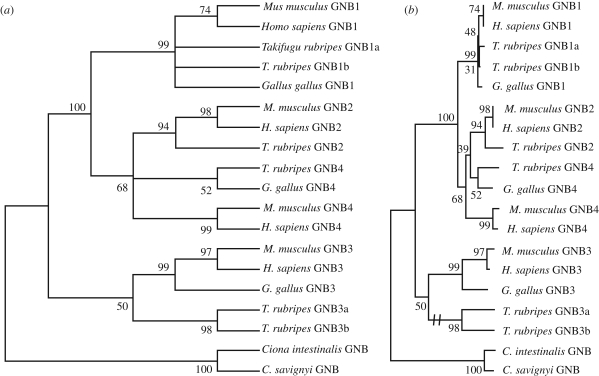
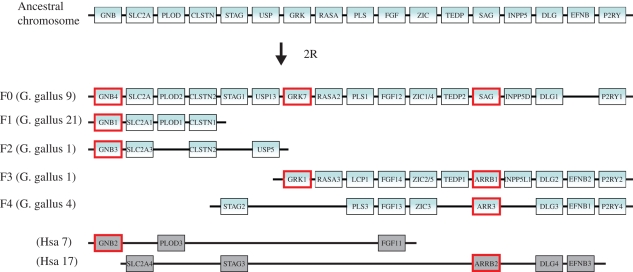
Vertebrate cones and rods in several cases use separate but related components for their signal transduction (opsins, G-proteins, ion channels, etc.). Some of these proteins are also used differentially in other cell types in the retina. Because cones, rods and other retinal cell types originated in early vertebrate evolution, it is of interest to see if their specific genes arose in the extensive gene duplications that took place in the ancestor of the jawed vertebrates (gnathostomes) by two tetraploidizations (genome doublings). The ancestor of teleost fishes subsequently underwent a third tetraploidization. Our previously reported analyses showed that several gene families in the vertebrate visual phototransduction cascade received new members in the basal tetraploidizations. We here expand these data with studies of additional gene families and vertebrate species. We conclude that no less than 10 of the 13 studied phototransduction gene families received additional members in the two basal vertebrate tetraploidizations. Also the remaining three families seem to have undergone duplications during the same time period but it is unclear if this happened as a result of the tetraploidizations. The implications of the many early vertebrate gene duplications for functional specialization of specific retinal cell types, particularly cones and rods, are discussed.
Figures

References
-
- Abi-Rached L., Gilles A., Shiina T., Pontarotti P., Inoko H.2002Evidence of en bloc duplication in vertebrate genomes. Nat. Genet. 31, 100–105 (doi:10.1038/ng855) - DOI - PubMed
-
- Ames J. B., Ikura M.2002Structure and membrane-targeting mechanism of retinal Ca2+-binding proteins, recoverin and GCAP-2. Adv. Exp. Med. Biol. 514, 333–348 - PubMed
-
- Applebury M. L., et al. 2000The murine cone photoreceptor: a single cone type expresses both S and M opsins with retinal spatial patterning. Neuron 27, 513–523 (doi:10.1016/S0896-6273(00)00062-3) - DOI - PubMed
-
- Arendt D.2008The evolution of cell types in animals: emerging principles from molecular studies. Nat. Rev. Genet. 9, 868–882 (doi:10.1038/nrg2416) - DOI - PubMed
-
- Balcueva E. A., Wang Q., Hughes H., Kunsch C., Yu Z., Robishaw J. D.2000Human G protein gamma(11) and gamma(14) subtypes define a new functional subclass. Exp. Cell Res. 257, 310–319 (doi:10.1006/excr.2000.4893) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources