

The evolution of early vertebrate photoreceptors
- PMID: 19720654
- PMCID: PMC2781863
- DOI: 10.1098/rstb.2009.0099
The evolution of early vertebrate photoreceptors
Abstract
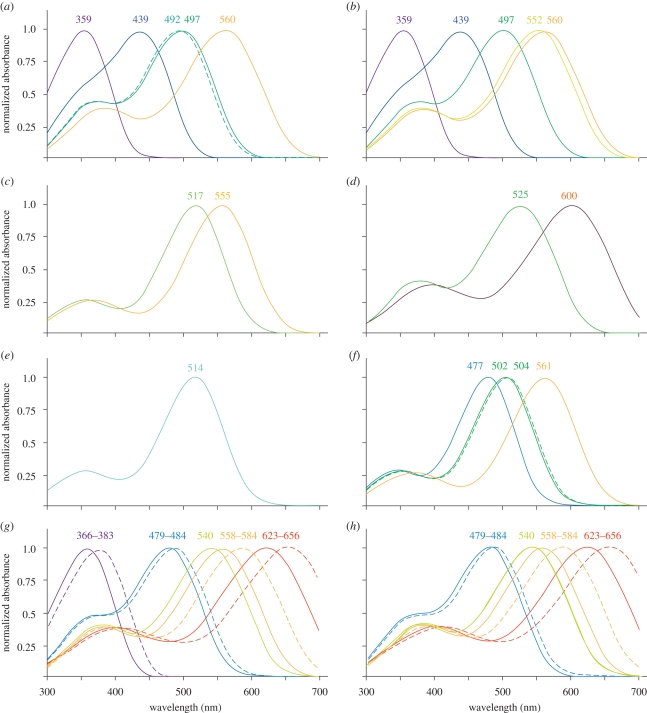
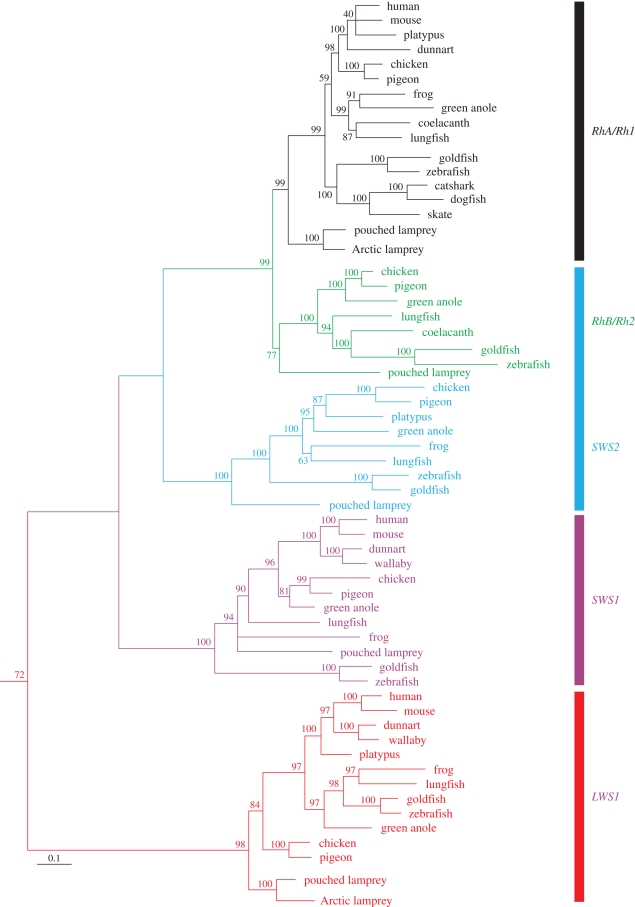
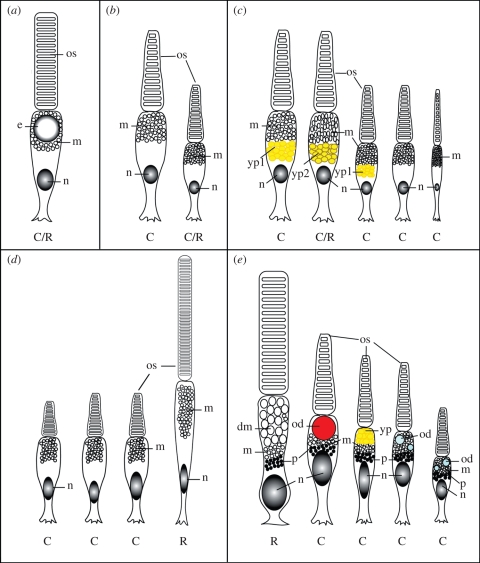
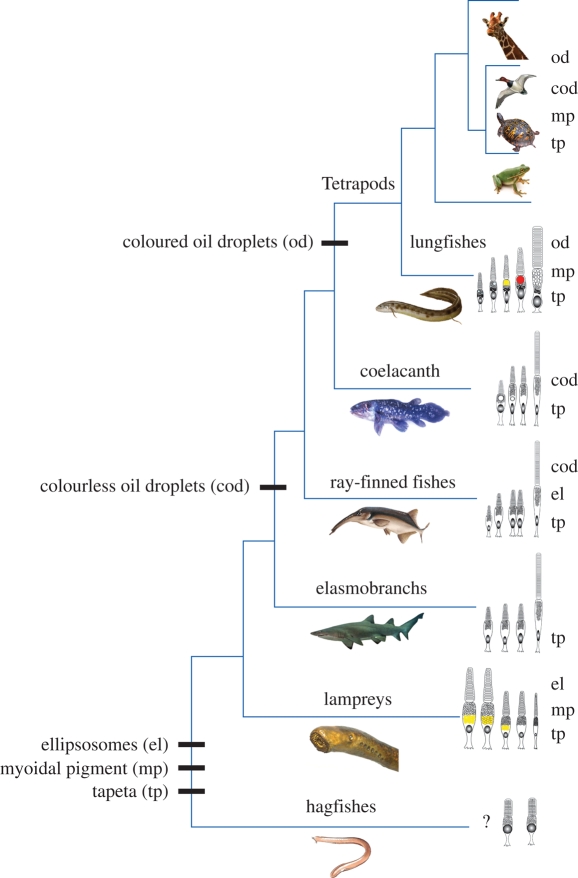
Meeting the challenge of sampling an ancient aquatic landscape by the early vertebrates was crucial to their survival and would establish a retinal bauplan to be used by all subsequent vertebrate descendents. Image-forming eyes were under tremendous selection pressure and the ability to identify suitable prey and detect potential predators was thought to be one of the major drivers of speciation in the Early Cambrian. Based on the fossil record, we know that hagfishes, lampreys, holocephalans, elasmobranchs and lungfishes occupy critical stages in vertebrate evolution, having remained relatively unchanged over hundreds of millions of years. Now using extant representatives of these 'living fossils', we are able to piece together the evolution of vertebrate photoreception. While photoreception in hagfishes appears to be based on light detection and controlling circadian rhythms, rather than image formation, the photoreceptors of lampreys fall into five distinct classes and represent a critical stage in the dichotomy of rods and cones. At least four types of retinal cones sample the visual environment in lampreys mediating photopic (and potentially colour) vision, a sampling strategy retained by lungfishes, some modern teleosts, reptiles and birds. Trichromacy is retained in cartilaginous fishes (at least in batoids and holocephalans), where it is predicted that true scotopic (dim light) vision evolved in the common ancestor of all living gnathostomes. The capacity to discriminate colour and balance the tradeoff between resolution and sensitivity in the early vertebrates was an important driver of eye evolution, where many of the ocular features evolved were retained as vertebrates progressed on to land.
Figures

References
-
- Arrese C. A., Hart N. S., Thomas N., Beazley L. D., Shand J.2002Trichromacy in Australian marsupials. Curr. Biol. 12, 657–660 (doi:10.1016/S0960-9822(02)00772-8) - DOI - PubMed
-
- Avery J. A., Bowmaker J. K.1982Visual pigments in the four-eyed fish. Anableps anableps. Nature 298, 62–64 (doi:10.1038/298062a0) - DOI
-
- Bailes H. J., Robinson S. R., Trezise A. E. O., Collin S. P.2006Morphology, characterisation and distribution of retinal photoreceptors in the Australian lungfish Neoceratodus forsteri (Krefft, 1870). J. Comp. Neurol. 494, 381–397 (doi:10.1002/cne.20809) - DOI - PubMed
-
- Bailes H. J., Davies W. L., Trezise A. E. O., Collin S. P.2007Visual pigments in a living fossil, the Australian lungfish Neoceratodus forsteri. BMC Evol. Biol. 7, 200 (doi:10.1186/1471–2148–7-200) - DOI - PMC - PubMed
-
- Barbour H. R., Archer M. A., Hart N. S., Thomas N., Dunlop S. A., Beazley L. D., Shand J.2002Retinal characteristics of the ornate dragon lizard, Ctenophorous ornatus. J. Comp. Neurol. 450, 334–344 (doi:10.1002/cne.10308) - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources