

Microbial NAD metabolism: lessons from comparative genomics
- PMID: 19721089
- PMCID: PMC2738131
- DOI: 10.1128/MMBR.00042-08
Microbial NAD metabolism: lessons from comparative genomics
Abstract
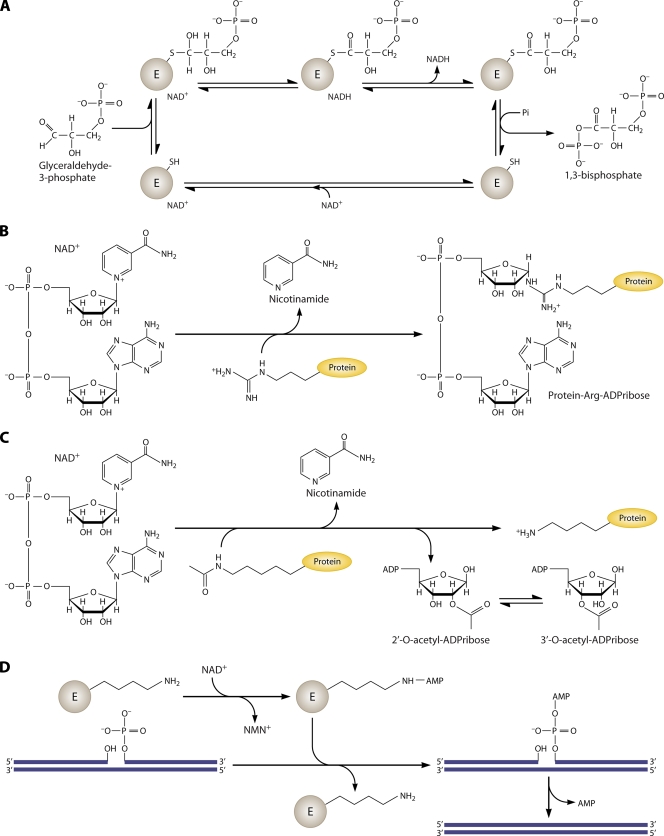
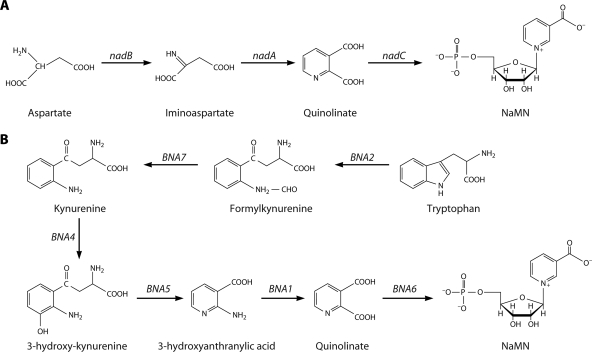
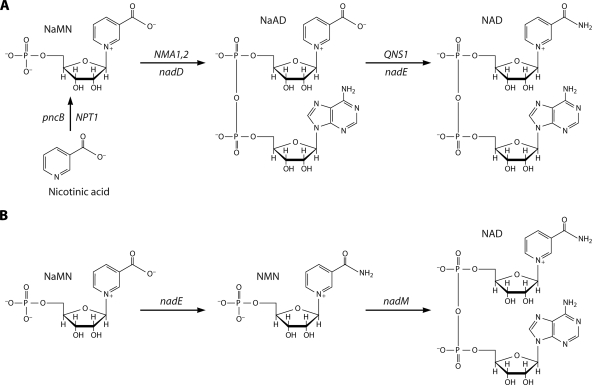
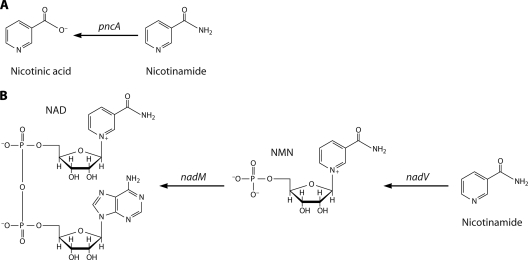
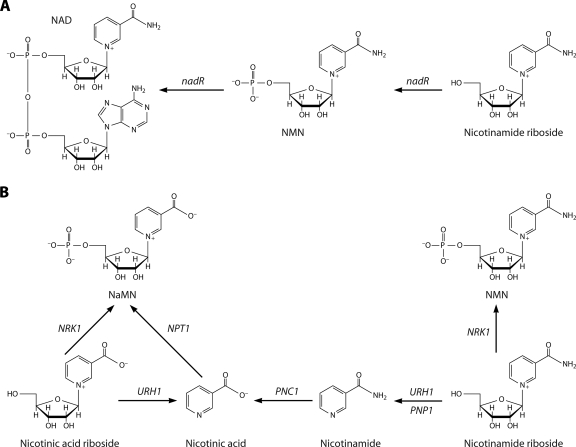
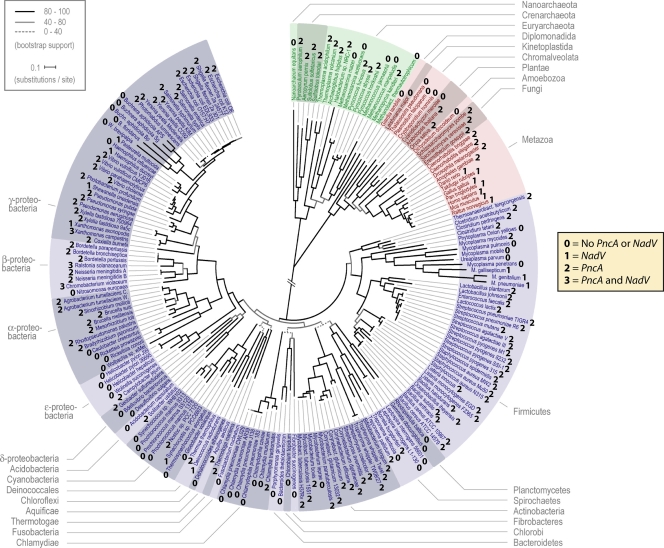
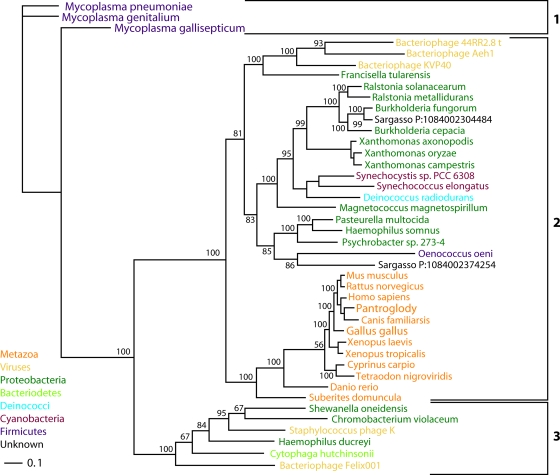
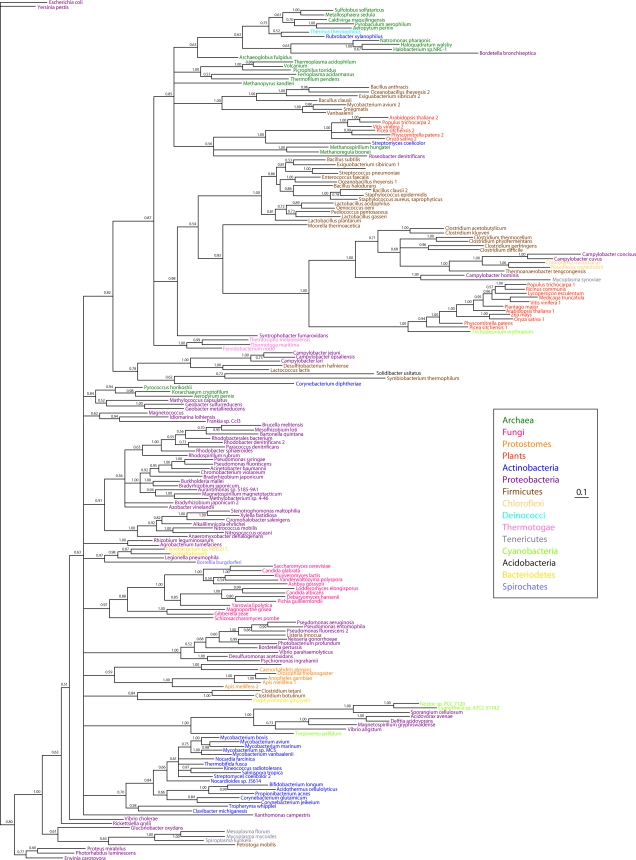
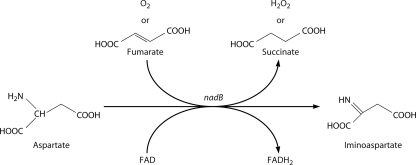
NAD is a coenzyme for redox reactions and a substrate of NAD-consuming enzymes, including ADP-ribose transferases, Sir2-related protein lysine deacetylases, and bacterial DNA ligases. Microorganisms that synthesize NAD from as few as one to as many as five of the six identified biosynthetic precursors have been identified. De novo NAD synthesis from aspartate or tryptophan is neither universal nor strictly aerobic. Salvage NAD synthesis from nicotinamide, nicotinic acid, nicotinamide riboside, and nicotinic acid riboside occurs via modules of different genes. Nicotinamide salvage genes nadV and pncA, found in distinct bacteria, appear to have spread throughout the tree of life via horizontal gene transfer. Biochemical, genetic, and genomic analyses have advanced to the point at which the precursors and pathways utilized by a microorganism can be predicted. Challenges remain in dissecting regulation of pathways.
Figures
References
-
- Anderson, R. M., K. J. Bitterman, J. G. Wood, O. Medvedik, H. Cohen, S. S. Lin, J. K. Manchester, J. I. Gordon, and D. A. Sinclair. 2002. Manipulation of a nuclear NAD+ salvage pathway delays aging without altering steady-state NAD+ levels. J. Biol. Chem. 27718881-18890. - PubMed
-
- Belenky, P., K. L. Bogan, and C. Brenner. 2007. NAD(+) metabolism in health and disease. Trends Biochem. Sci. 3212-19. - PubMed
-
- Belenky, P., F. G. Racette, K. L. Bogan, J. M. McClure, J. S. Smith, and C. Brenner. 2007. Nicotinamide riboside promotes Sir2 silencing and extends lifespan via Nrk and Urh1/Pnp1/Meu1 pathways to NAD(+). Cell 129473-484. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
