

Germline deletion of AMP-activated protein kinase beta subunits reduces bone mass without altering osteoclast differentiation or function
- PMID: 19723702
- PMCID: PMC2797037
- DOI: 10.1096/fj.09-137158
Germline deletion of AMP-activated protein kinase beta subunits reduces bone mass without altering osteoclast differentiation or function
Abstract
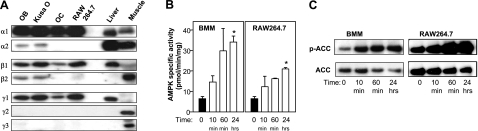
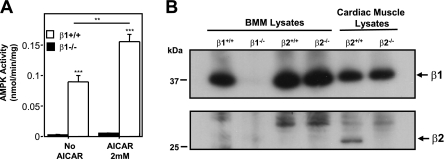
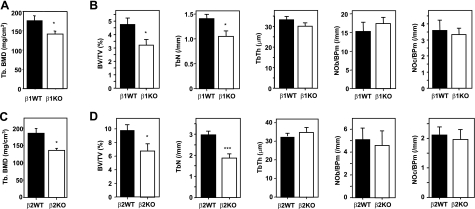
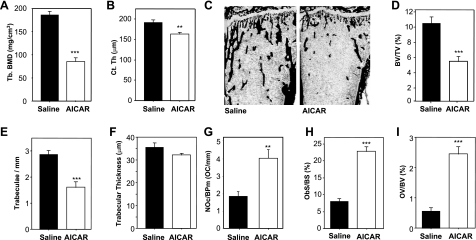
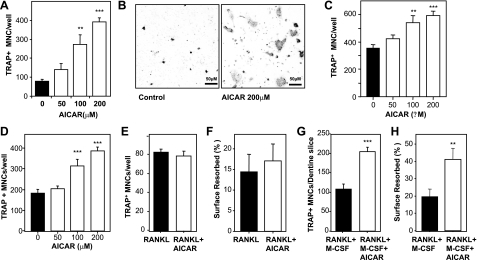
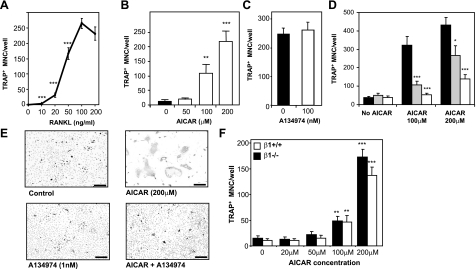
Since AMP-activated protein kinase (AMPK) plays important roles in modulating metabolism in response to diet and exercise, both of which influence bone mass, we examined the influence of AMPK on bone mass in mice. AMPK is an alphabetagamma heterotrimer where the beta subunit anchors the alpha catalytic and gamma regulatory subunits. Germline deletion of either AMPK beta1 or beta2 subunit isoforms resulted in reduced trabecular bone density and mass, but without effects on osteoclast (OC) or osteoblast (OB) numbers, as compared to wild-type littermate controls. We tested whether activating AMPK in vivo would enhance bone density but found AICA-riboside treatment caused a profound loss of trabecular bone volume (49.5%) and density and associated increased OC numbers. Consistent with this, AICA-riboside strongly stimulated OC differentiation in vitro, in an adenosine kinase-dependent manner. OCs and macrophages (unlike OBs) lacked AMPK beta2 subunit expression, and when generated from AMPK beta1(-/-) mice displayed no detectable AMPK activity. Nevertheless, AICA-riboside was equally effective at stimulating OC differentiation from wild-type or beta1(-/-) progenitors, indicating that AMPK is not essential for OC differentiation or the stimulatory action of AICA-riboside. These results show that AMPK is required to maintain normal bone density, but not through bone cell differentiation, and does not mediate powerful osteolytic effects of AICA-riboside.
Figures
References
-
- Karsenty G. Convergence between bone and energy homeostases: leptin regulation of bone mass. Cell Metab. 2006;4:341–348. - PubMed
-
- Seibel M J. Nutrition and molecular markers of bone remodelling. Curr Opin Clin Nutr Metab Care. 2002;5:525–531. - PubMed
-
- Scott J W, Oakhill J S, van Denderen B J W. AMPK/SNF1 structure: a ménage a trois of energy-sensing. Front Biosci. 2009;14:596–610. - PubMed
-
- Sun W, Lee T S, Zhu M, Gu C, Wang Y, Zhu Y, Shyy J Y. Statins activate AMP-activated protein kinase in vitro and in vivo. Circulation. 2006;114:2655–2662. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
