

Antiapoptotic roles of ceramide-synthase-6-generated C16-ceramide via selective regulation of the ATF6/CHOP arm of ER-stress-response pathways
- PMID: 19723703
- PMCID: PMC2797032
- DOI: 10.1096/fj.09-135087
Antiapoptotic roles of ceramide-synthase-6-generated C16-ceramide via selective regulation of the ATF6/CHOP arm of ER-stress-response pathways
Abstract
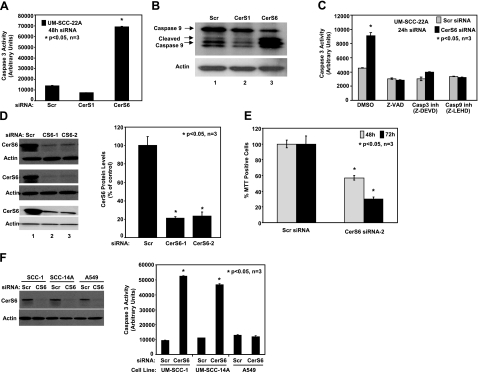
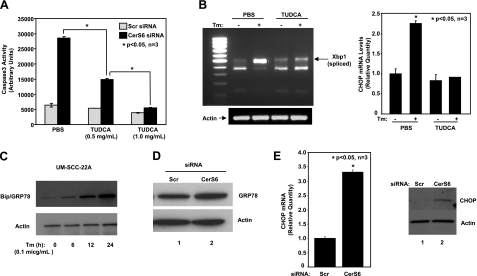
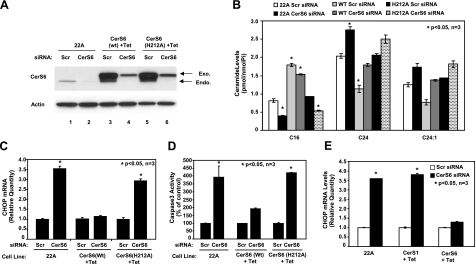
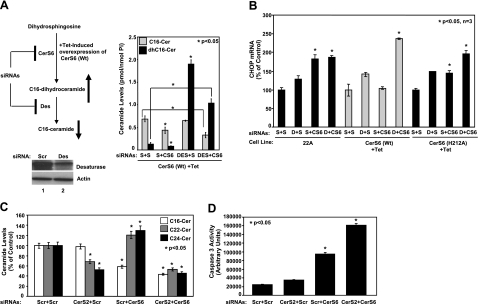
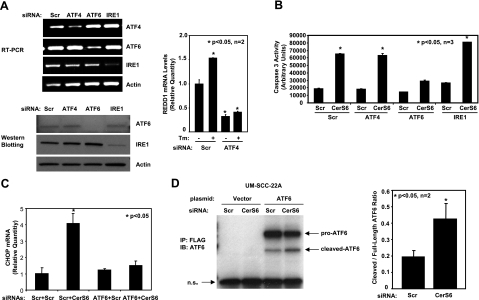
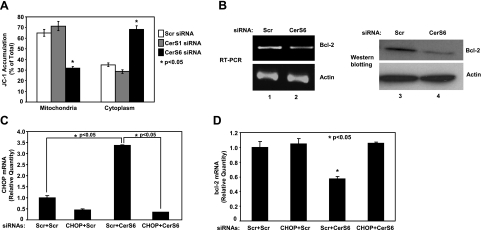
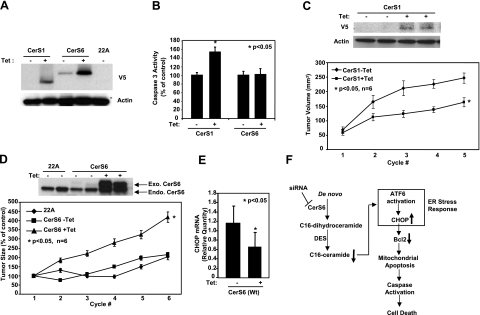
Emerging results suggest that ceramides with different fatty acid chain lengths might play distinct functions in the regulation of tumor growth and therapy. Here we report that de novo-generated C(18)- and C(16)-ceramides by ceramide synthases 1 and 6 (CerS1 and CerS6) play opposing proapoptotic and prosurvival roles, respectively, in human head and neck squamous cell carcinomas (HNSCCs). Unexpectedly, knockdown of CerS6/C(16)-ceramide using small interfering RNA induced endoplasmic reticulum (ER)-stress-mediated apoptosis. Reconstitution of C(16)-ceramide generation by induced expression of wild-type CerS6, but not its catalytically inactive mutant, protected cells from cell death induced by knockdown of CerS6. Moreover, using molecular tools coupled with analysis of sphingolipid metabolism showed that generation of C(16)-ceramide, and not dihydro-C(16)-ceramide, by induced expression of CerS6 rescued cells from ER stress and apoptosis. Mechanistically, regulation of ER-stress-induced apoptosis by CerS6/C(16)-ceramide was linked to the activation of a specific arm, ATF6/CHOP, of the unfolded protein response pathway. Notably, while expression of CerS1/C(18)-ceramide inhibited HNSCC xenograft growth, CerS6/C(16)-ceramide significantly protected ER stress, leading to enhanced tumor development and growth in vivo, consistent with their pro- and antiapoptotic roles, respectively. Thus, these data reveal an unexpected and novel prosurvival role of CerS6/C(16)-ceramide involved in the protection against ER-stress-induced apoptosis and induction of HNSCC tumor growth.
Figures
References
-
- Ogretmen B, Hannun Y A. Biologically active sphingolipids in cancer pathogenesis and treatment. Nat Rev Cancer. 2004;4:604–616. - PubMed
-
- Mehta S, Blackinton D, Omar I, Kouttab N, Myrick D, Klostergaard J, Wanebo H. Combined cytotoxic action of paclitaxel and ceramide against the human Tu138 head and neck squamous carcinoma cell line. Cancer Chemother Pharmacol. 2000;46:85–92. - PubMed
-
- Senkal C E, Ponnusamy S, Rossi M J, Bialewski J, Sinha D, Jiang J C, Jazwinski S M, Hannun Y A, Ogretmen B. Role of human longevity assurance gene 1 and C18-ceramide in chemotherapy-induced cell death in human head and neck squamous cell carcinomas. Mol Cancer Ther. 2007;6:712–722. - PubMed
-
- Swanton C, Marani M, Pardo O, Warne P H, Kelly G, Sahai E, Elustondo F, Chang J, Temple J, Ahmed A A, Brenton J D, Downward J, Nicke B. Regulators of mitotic arrest and ceramide metabolism are determinants of sensitivity to paclitaxel and other chemotherapeutic drugs. Cancer Cell. 2007;11:498–512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
