

Nucleosome assembly proteins bind to Epstein-Barr virus nuclear antigen 1 and affect its functions in DNA replication and transcriptional activation
- PMID: 19726498
- PMCID: PMC2772708
- DOI: 10.1128/JVI.00931-09
Nucleosome assembly proteins bind to Epstein-Barr virus nuclear antigen 1 and affect its functions in DNA replication and transcriptional activation
Abstract
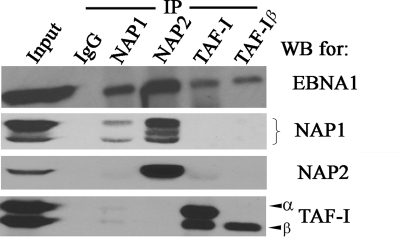
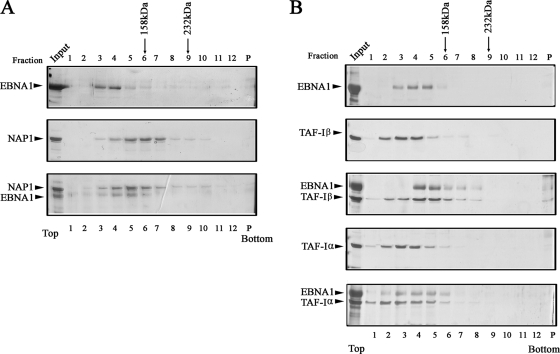
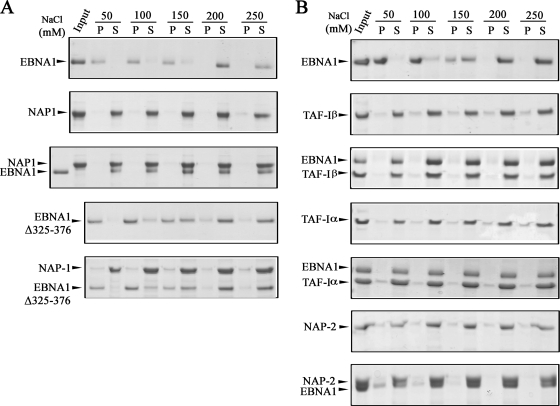
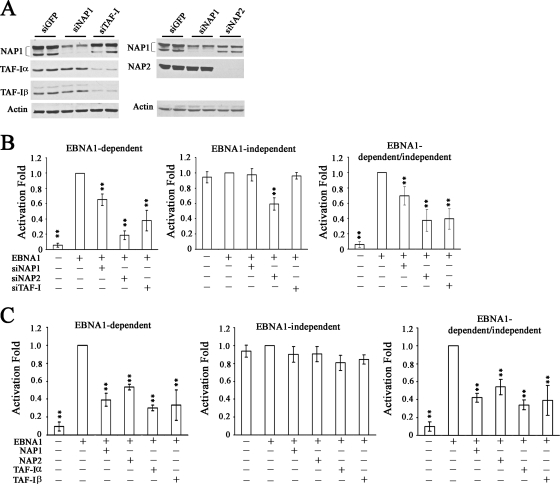
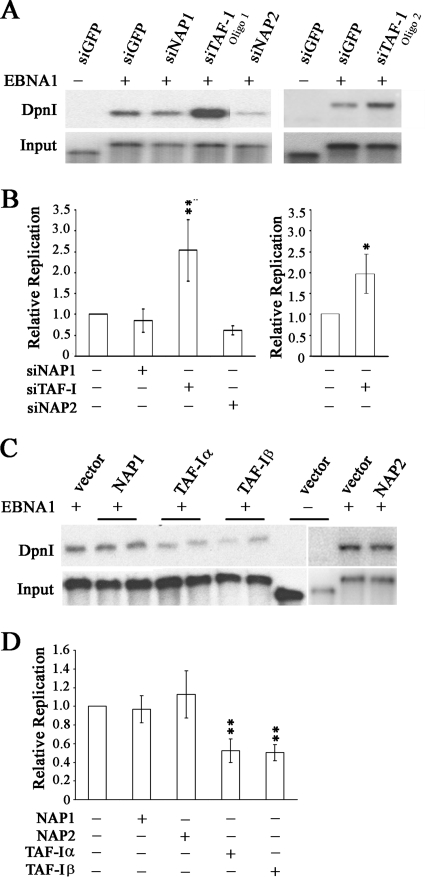
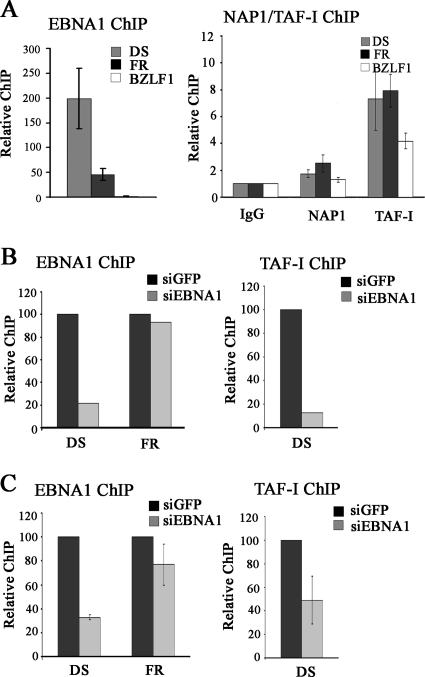
The EBNA1 protein of Epstein-Barr virus (EBV) plays several important roles in EBV latent infection, including activating DNA replication from the latent origin of replication (oriP) and activating the transcription of other latency genes within the EBV chromatin. These functions require EBNA1 binding to the DS and FR elements within oriP, respectively, although how these interactions activate these processes is not clear. We previously identified interactions of EBNA1 with the related nucleosome assembly proteins NAP1 and TAF-I, known to affect the replication and transcription of other chromatinized templates. We have further investigated these interactions, showing that EBNA1 binds directly to NAP1 and to the beta isoform of TAF-I (also called SET) and that these interactions greatly increase the solubility of EBNA1 in vitro. These interactions were confirmed in EBV-infected cells, and chromatin immunoprecipitation with these cells showed that NAP1 and TAF-I both localized with EBNA1 to the FR element, while only TAF-I was detected with EBNA1 at the DS element. In keeping with these observations, alteration of the NAP1 or TAF-Ibeta level by RNA interference and overexpression inhibited transcriptional activation by EBNA1 in FR reporter assays. In addition, EBNA1-mediated DNA replication was stimulated when TAF-I (but not NAP1) was downregulated and was inhibited by TAF-Ibeta overexpression. The results indicate that the interaction of EBNA1 with NAP1 and TAF-I is important for transcriptional activation and that EBNA1 recruits TAF-I to the DS element, where it negatively regulates DNA replication.
Figures
Similar articles
-
EBNA1 efficiently assembles on chromatin containing the Epstein-Barr virus latent origin of replication.Virology. 2003 Oct 25;315(2):398-408. doi: 10.1016/s0042-6822(03)00561-0. Virology. 2003. PMID: 14585343
-
Structural Basis for Cooperative Binding of EBNA1 to the Epstein-Barr Virus Dyad Symmetry Minimal Origin of Replication.J Virol. 2019 Sep 30;93(20):e00487-19. doi: 10.1128/JVI.00487-19. Print 2019 Oct 15. J Virol. 2019. PMID: 31142669 Free PMC article.
-
EBNA1-mediated recruitment of a histone H2B deubiquitylating complex to the Epstein-Barr virus latent origin of DNA replication.PLoS Pathog. 2009 Oct;5(10):e1000624. doi: 10.1371/journal.ppat.1000624. Epub 2009 Oct 16. PLoS Pathog. 2009. PMID: 19834552 Free PMC article.
-
Replication licensing of the EBV oriP minichromosome.Curr Top Microbiol Immunol. 2001;258:13-33. doi: 10.1007/978-3-642-56515-1_2. Curr Top Microbiol Immunol. 2001. PMID: 11443858 Review.
-
EBNA1 and host factors in Epstein-Barr virus latent DNA replication.Curr Opin Virol. 2012 Dec;2(6):733-9. doi: 10.1016/j.coviro.2012.09.005. Epub 2012 Sep 30. Curr Opin Virol. 2012. PMID: 23031715 Review.
Cited by
-
B Cell-Specific Transcription Activator PAX5 Recruits p300 To Support EBNA1-Driven Transcription.J Virol. 2020 Mar 17;94(7):e02028-19. doi: 10.1128/JVI.02028-19. Print 2020 Mar 17. J Virol. 2020. PMID: 31941781 Free PMC article.
-
Epstein-Barr virus EBNA1 protein regulates viral latency through effects on let-7 microRNA and dicer.J Virol. 2014 Oct;88(19):11166-77. doi: 10.1128/JVI.01785-14. Epub 2014 Jul 16. J Virol. 2014. PMID: 25031339 Free PMC article.
-
Opportunities to Target the Life Cycle of Epstein-Barr Virus (EBV) in EBV-Associated Lymphoproliferative Disorders.Front Oncol. 2019 Mar 15;9:127. doi: 10.3389/fonc.2019.00127. eCollection 2019. Front Oncol. 2019. PMID: 30931253 Free PMC article. Review.
-
In Vitro Efficacy of PEI-Derived Lipopolymers in Silencing of Toxic Proteins in a Neuronal Model of Huntington's Disease.Pharmaceutics. 2025 May 30;17(6):726. doi: 10.3390/pharmaceutics17060726. Pharmaceutics. 2025. PMID: 40574039 Free PMC article.
-
Cis and trans acting factors involved in human cytomegalovirus experimental and natural latent infection of CD14 (+) monocytes and CD34 (+) cells.PLoS Pathog. 2013;9(5):e1003366. doi: 10.1371/journal.ppat.1003366. Epub 2013 May 23. PLoS Pathog. 2013. PMID: 23717203 Free PMC article. Clinical Trial.
References
-
- Abu-Daya, A., W. M. Steer, A. F. Trollope, C. E. Friedeberg, R. K. Patient, A. W. Thorne, and M. J. Guille. 2005. Zygotic nucleosome assembly protein-like 1 has a specific, non-cell autonomous role in hematopoiesis. Blood 106:514-520. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
