

Receptor binding and low pH coactivate oncogenic retrovirus envelope-mediated fusion
- PMID: 19726505
- PMCID: PMC2772678
- DOI: 10.1128/JVI.00748-09
Receptor binding and low pH coactivate oncogenic retrovirus envelope-mediated fusion
Abstract
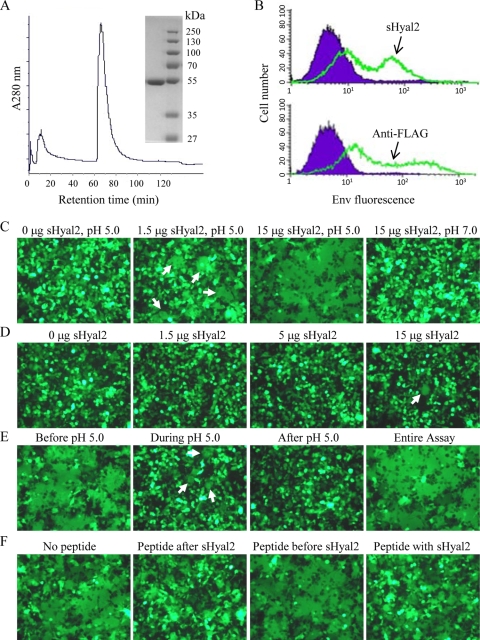
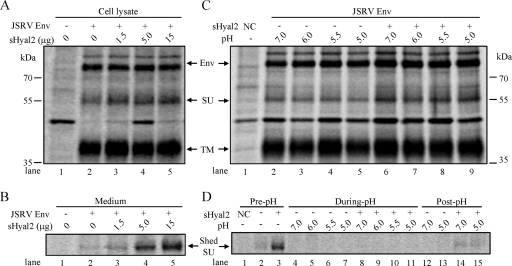
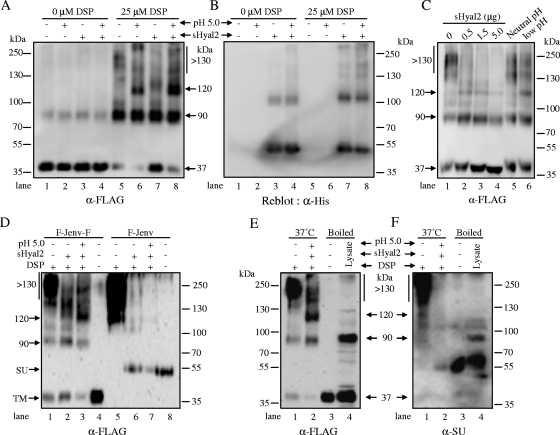
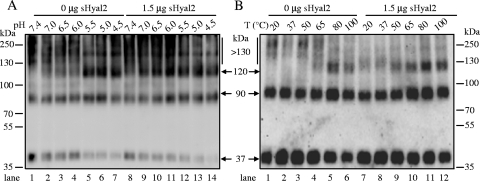
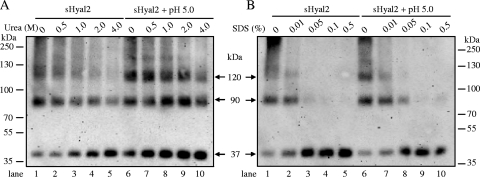
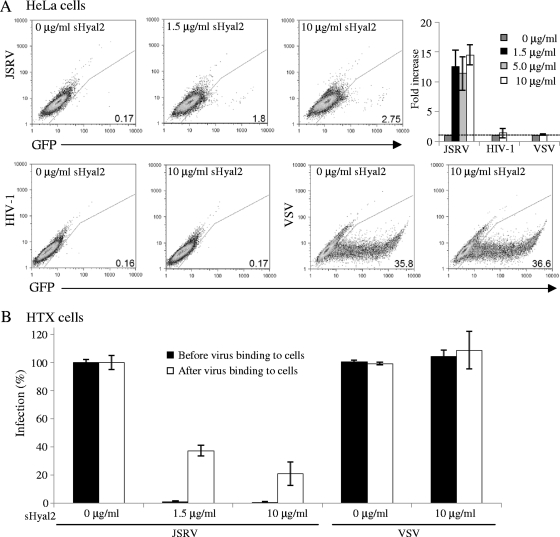
Fusion of enveloped viruses with host cells is triggered by either receptor binding or low pH but rarely requires both except for avian sarcoma leukosis virus (ASLV). We recently reported that membrane fusion mediated by an oncogenic Jaagsiekte sheep retrovirus (JSRV) envelope (Env) requires an acidic pH, yet receptor overexpression is required for this process to occur. Here we show that a soluble form of the JSRV receptor, sHyal2, promoted JSRV Env-mediated fusion at a low pH in normally fusion-negative cells and that this effect was blocked by a synthetic peptide analogous to the C-terminal heptad repeat of JSRV Env. In contrast to the receptor of ASLV, sHyal2 induced pronounced shedding of the JSRV surface subunit, as well as unstable conformational rearrangement of its transmembrane (TM) subunit, yet full activation of JSRV Env fusogenicity, associated with strong TM oligomerization, required both sHyal2 and low pH. Consistently, sHyal2 enabled transduction of nonpermissive cells by JSRV Env pseudovirions, with low efficiency, but substantially blocked viral entry into permissive cells at both binding and postbinding steps, indicating that sHyal2 prematurely activates JSRV Env-mediated fusion. Altogether, our study supports a model that receptor priming promotes fusion activation of JSRV Env at a low pH, and that the underlying mechanism is likely to be different from that of ASLV. Thus, JSRV may provide a useful alternate model for the better understanding of virus fusion and cell entry.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
