

C-terminal phenylalanine of bacteriophage T7 single-stranded DNA-binding protein is essential for strand displacement synthesis by T7 DNA polymerase at a nick in DNA
- PMID: 19726688
- PMCID: PMC2781589
- DOI: 10.1074/jbc.M109.024059
C-terminal phenylalanine of bacteriophage T7 single-stranded DNA-binding protein is essential for strand displacement synthesis by T7 DNA polymerase at a nick in DNA
Abstract
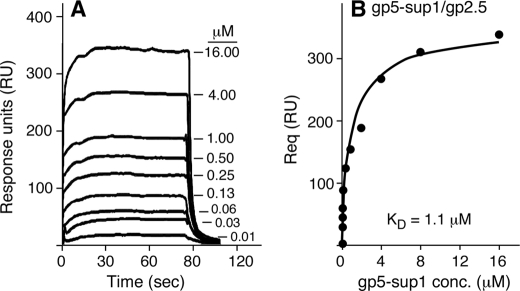
Single-stranded DNA-binding protein (gp2.5), encoded by gene 2.5 of bacteriophage T7, plays an essential role in DNA replication. Not only does it remove impediments of secondary structure in the DNA, it also modulates the activities of the other replication proteins. The acidic C-terminal tail of gp2.5, bearing a C-terminal phenylalanine, physically and functionally interacts with the helicase and DNA polymerase. Deletion of the phenylalanine or substitution with a nonaromatic amino acid gives rise to a dominant lethal phenotype, and the altered gp2.5 has reduced affinity for T7 DNA polymerase. Suppressors of the dominant lethal phenotype have led to the identification of mutations in gene 5 that encodes the T7 DNA polymerase. The altered residues in the polymerase are solvent-exposed and lie in regions that are adjacent to the bound DNA. gp2.5 lacking the C-terminal phenylalanine has a lower affinity for gp5-thioredoxin relative to the wild-type gp2.5, and this affinity is partially restored by the suppressor mutations in DNA polymerase. gp2.5 enables T7 DNA polymerase to catalyze strand displacement DNA synthesis at a nick in DNA. The resulting 5'-single-stranded DNA tail provides a loading site for T7 DNA helicase. gp2.5 lacking the C-terminal phenylalanine does not support this event with wild-type DNA polymerase but does to a limited extent with T7 DNA polymerase harboring the suppressor mutations.
Figures






Similar articles
-
Gp2.5, the multifunctional bacteriophage T7 single-stranded DNA binding protein.Semin Cell Dev Biol. 2019 Feb;86:92-101. doi: 10.1016/j.semcdb.2018.03.018. Epub 2018 Mar 28. Semin Cell Dev Biol. 2019. PMID: 29588157 Free PMC article. Review.
-
Mutations in the gene 5 DNA polymerase of bacteriophage T7 suppress the dominant lethal phenotype of gene 2.5 ssDNA binding protein lacking the C-terminal phenylalanine.Mol Microbiol. 2009 May;72(4):869-80. doi: 10.1111/j.1365-2958.2009.06682.x. Epub 2009 Mar 30. Mol Microbiol. 2009. PMID: 19400798
-
The carboxyl-terminal domain of bacteriophage T7 single-stranded DNA-binding protein modulates DNA binding and interaction with T7 DNA polymerase.J Biol Chem. 2003 Aug 8;278(32):29538-45. doi: 10.1074/jbc.M304318200. Epub 2003 May 24. J Biol Chem. 2003. PMID: 12766155
-
Acidic carboxyl-terminal domain of gene 2.5 protein of bacteriophage T7 is essential for protein-protein interactions.J Biol Chem. 1994 Feb 18;269(7):5270-8. J Biol Chem. 1994. PMID: 8106511
-
The Replication System of Bacteriophage T7.Enzymes. 2016;39:89-136. doi: 10.1016/bs.enz.2016.02.001. Epub 2016 Mar 28. Enzymes. 2016. PMID: 27241928 Review.
Cited by
-
Catalytically inactive T7 DNA polymerase imposes a lethal replication roadblock.J Biol Chem. 2020 Jul 10;295(28):9542-9550. doi: 10.1074/jbc.RA120.013738. Epub 2020 May 19. J Biol Chem. 2020. PMID: 32430399 Free PMC article.
-
Flap endonuclease of bacteriophage T7: Possible roles in RNA primer removal, recombination and host DNA breakdown.Bacteriophage. 2014 Mar 11;4:e28507. doi: 10.4161/bact.28507. eCollection 2014. Bacteriophage. 2014. PMID: 25105057 Free PMC article.
-
Choreography of bacteriophage T7 DNA replication.Curr Opin Chem Biol. 2011 Oct;15(5):580-6. doi: 10.1016/j.cbpa.2011.07.024. Epub 2011 Sep 9. Curr Opin Chem Biol. 2011. PMID: 21907611 Free PMC article. Review.
-
Gp2.5, the multifunctional bacteriophage T7 single-stranded DNA binding protein.Semin Cell Dev Biol. 2019 Feb;86:92-101. doi: 10.1016/j.semcdb.2018.03.018. Epub 2018 Mar 28. Semin Cell Dev Biol. 2019. PMID: 29588157 Free PMC article. Review.
-
Mechanisms of mutagenesis: DNA replication in the presence of DNA damage.Mutat Res Rev Mutat Res. 2016 Apr-Jun;768:53-67. doi: 10.1016/j.mrrev.2016.03.006. Epub 2016 Apr 7. Mutat Res Rev Mutat Res. 2016. PMID: 27234563 Free PMC article. Review.
References
-
- Chase J. W., Williams K. R. (1986) Annu. Rev. Biochem. 55, 103–136 - PubMed
-
- Benkovic S. J., Valentine A. M., Salinas F. (2001) Annu. Rev. Biochem. 70, 181–208 - PubMed
-
- O'Donnell M. (2006) J. Biol. Chem. 281, 10653–10656 - PubMed
-
- Hamdan S. M., Richardson C. C. (2009) Annu. Rev. Biochem. 78, 205–243 - PubMed
-
- Lohman T. M., Ferrari M. E. (1994) Annu. Rev. Biochem. 63, 527–570 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
