

Myofibril-inducing RNA (MIR) is essential for tropomyosin expression and myofibrillogenesis in axolotl hearts
- PMID: 19728883
- PMCID: PMC2752452
- DOI: 10.1186/1423-0127-16-81
Myofibril-inducing RNA (MIR) is essential for tropomyosin expression and myofibrillogenesis in axolotl hearts
Abstract
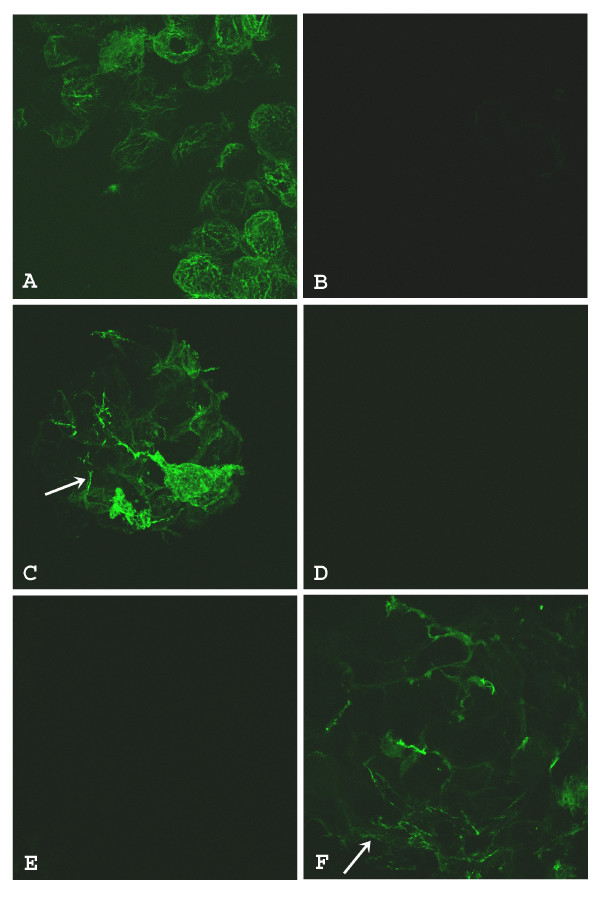
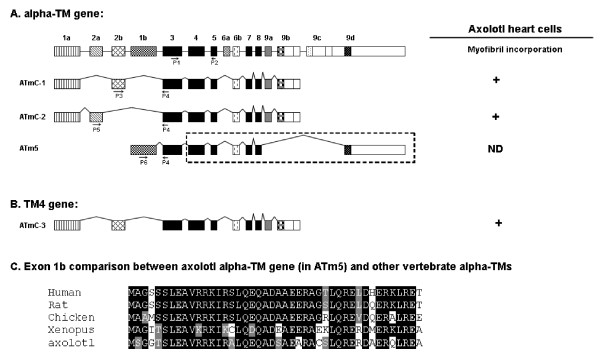
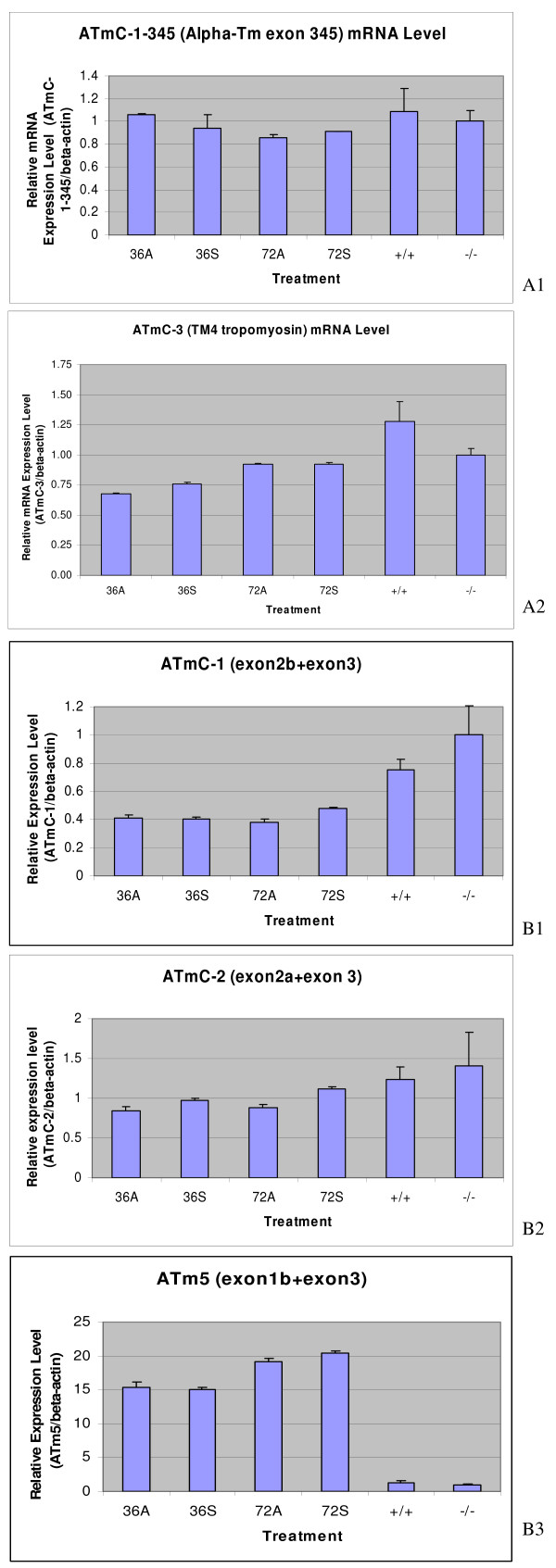
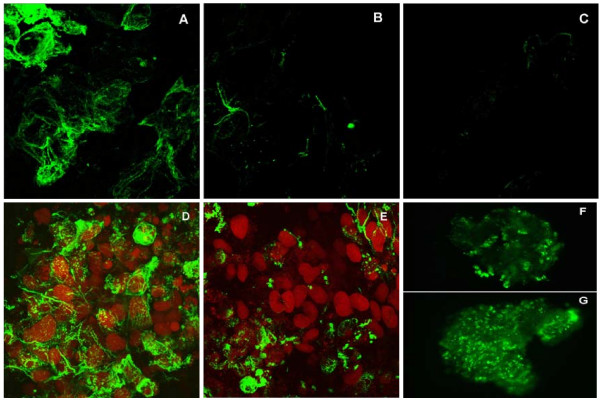
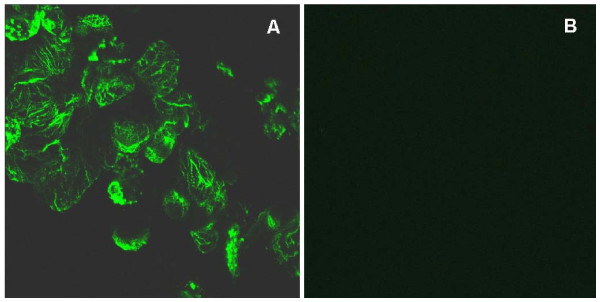
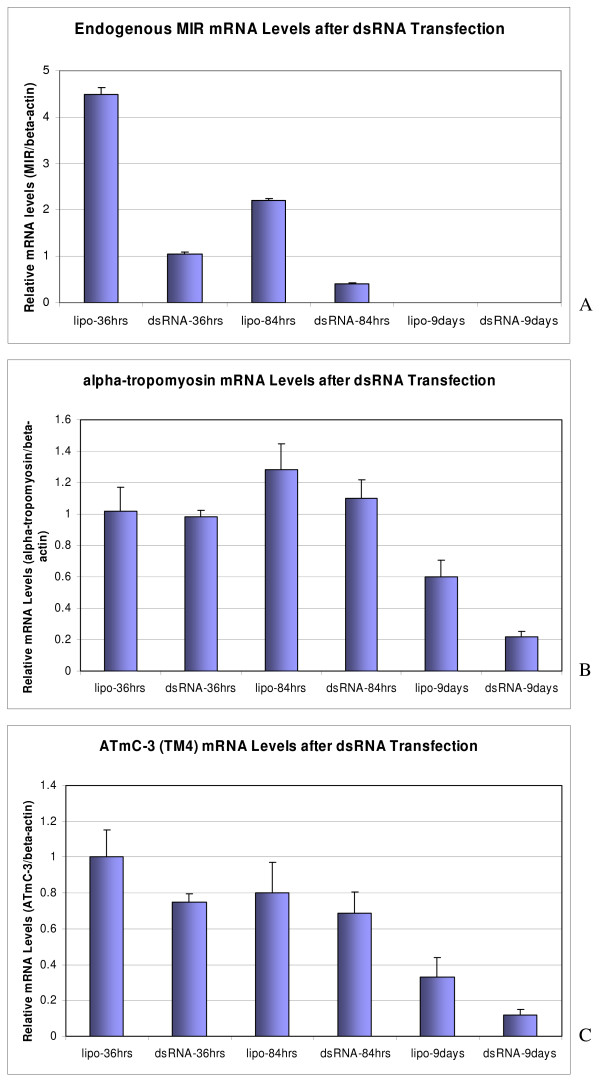
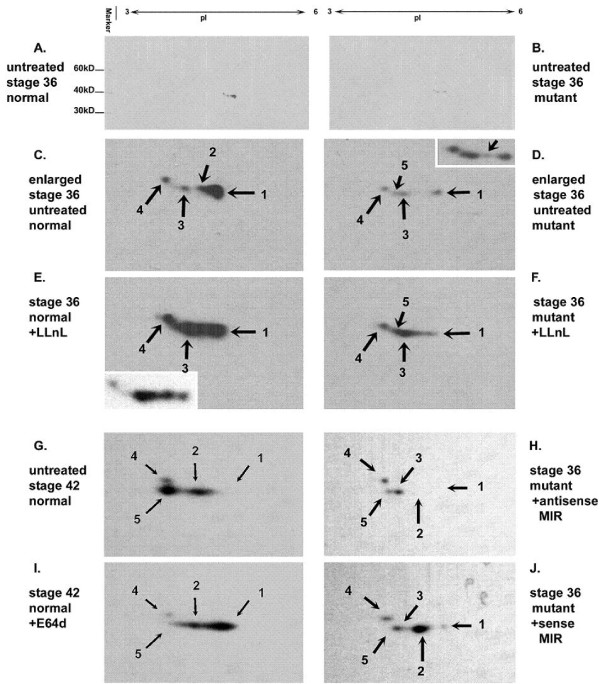
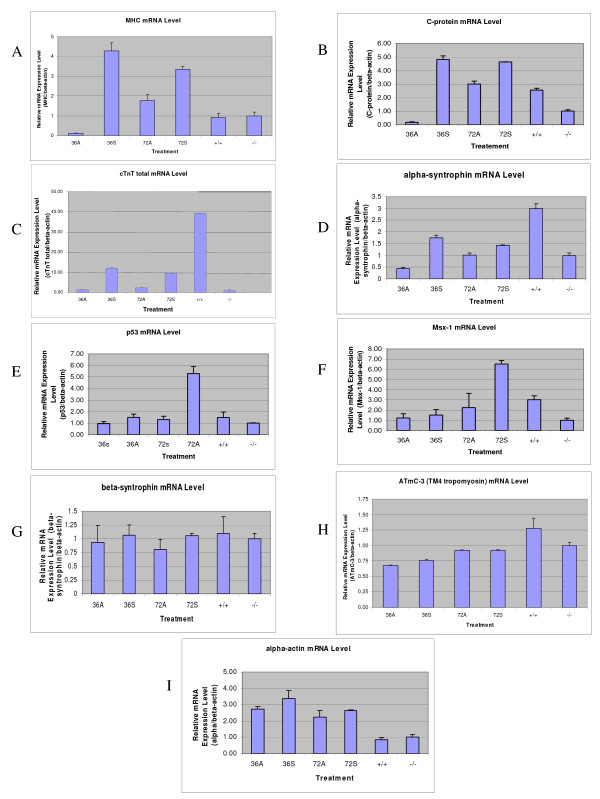
The Mexican axolotl, Ambystoma mexicanum, carries the naturally-occurring recessive mutant gene 'c' that results in a failure of homozygous (c/c) embryos to form hearts that beat because of an absence of organized myofibrils. Our previous studies have shown that a noncoding RNA, Myofibril-Inducing RNA (MIR), is capable of promoting myofibrillogenesis and heart beating in the mutant (c/c) axolotls. The present study demonstrates that the MIR gene is essential for tropomyosin (TM) expression in axolotl hearts during development. Gene expression studies show that mRNA expression of various tropomyosin isoforms in untreated mutant hearts and in normal hearts knocked down with double-stranded MIR (dsMIR) are similar to untreated normal. However, at the protein level, selected tropomyosin isoforms are significantly reduced in mutant and dsMIR treated normal hearts. These results suggest that MIR is involved in controlling the translation or post-translation of various TM isoforms and subsequently of regulating cardiac contractility.
Figures

References
-
- LaFrance SM, Lemanski LF. Imunofluorescent confocal analysis of tropomyosin in developing hearts of normal and cardiac mutant axolotls. Int J Devel Biol. 1994;38:695–700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
