

Review
doi: 10.1016/j.tibs.2009.05.003.
Epub 2009 Sep 2.
Re-capping the message
Affiliations
- PMID: 19729311
- PMCID: PMC2743798
- DOI: 10.1016/j.tibs.2009.05.003
Item in Clipboard
Review
Re-capping the message
Trends Biochem Sci.
2009 Sep.
Abstract
The 5'-cap structure that typifies all polymerase II-transcribed RNAs plays important roles in pre-mRNA processing and mRNA export, translation and quality control. Removal of the cap is a regulated process that is considered to be the first irreversible step in mRNA decay. An emerging view challenges this idea: mRNAs have been identified in mammalian cells that lack sequences from their 5' ends but nevertheless appear to be modified with a cap or cap-like structure. Furthermore, a cytoplasmic form of capping enzyme was recently identified that, together with a novel kinase, generates capped ends from cleaved RNAs. These and other findings provide evidence for re-capping and its possible functions.
Figures
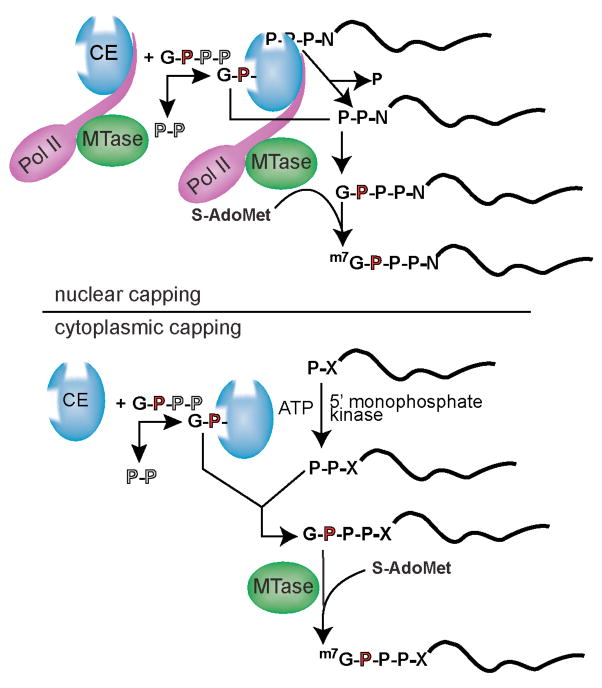
Nuclear capping occurs co-transcriptionally on all RNA polymerase II-synthesized RNA, and recent evidence indicates that cytoplasmic capping can occur by a related mechanism. (i) The first steps of nuclear capping. Binding to the C-terminal domain of the large subunit of RNA polymerase II (Pol II; purple) positions the nuclear capping enzyme (CE; blue) near the 5′ end of newly synthesized transcripts. The N-terminal triphosphatase domain of capping enzyme hydrolyzes the 5′-triphosphate end of the first transcribed nucleotide (P-P-P-N, where N denotes the first transcribed nucleotide) to a diphosphate (P-P-N) capping substrate. Capping enzyme then transfers GMP that is bound covalently at its C-terminal guanylyltransferase domain to create the 5′-G(5′)-P-P-P-(5′)N terminus (G-P-P-P-N). RNA (guanine-7)-methytransferase (MTase; green) is also positioned near the transcript 5′ end via binding to Pol II. Transfer of a methyl group from S-adenosylmethionine (S-AdoMet) onto the N7 position of the terminal guanine generates 5′-m7G(5′)-P-P-P-(5′)N, thereby completing the synthesis of the cap structure referred to as cap 0. (ii) The steps of cytoplasmic capping. As in nuclear capping, GMP is bound covalently to the guanylyltransferase domain of capping enzyme. However, in contrast to nuclear capping, cytoplasmic capping enzyme forms a complex with a 5′-monophosphate kinase that converts uncapped RNA having a 5′-monophosphate (P-X) end to a diphosphate capping substrate (P-P-X). As in nuclear capping, GMP that is bound covalently to the guanylyltransferase domain of capping enzyme is then transferred to create the G(5′)-P-P-P-(5′)N end (G-P-P-P-X). Although it is present in the cytoplasm, cap methyltransferase (MTase; green) is not part of the cytoplasmic capping enzyme complex. The process by which the cap is methylated during cytoplasmic capping remains undetermined.

mRNA that is actively engaged in translation is bound by eIF4E (4E; beige) and eIF4G (4G; blue) at its 5′ end, poly(A)-binding protein (not shown for simplicity) at its 3′-poly(A) tail, and 80S ribosomes (green circles), among many other factors. The poly(A) tail is shortened by one or more deadenylases until a limit length of ~30 residues is achieved (Aoligo), at which point ribosomes no longer initiate translation, translationally active ribosomes presumably dissociate after directing translation termination, and the mRNA transits into a translationally inactive state. This transition, which can result in microscopically detectable P bodies, is likely facilitated by translational inhibitors, including Pat1 (red), the p54 (also called Rck) helicase (yellow) and, possibly, microRNAs. Little is known about the order of assembly, but a variety of proteins are recruited that include the decapping activators Lsm1-7 (blue ring), enhancer of mRNA decapping 3 homologue (Edc3; brown), Hedls (orange) and GW182 (blue oblong), the decapping enzyme Dcp2 (dark pink) and its activator Dcp1 (light pink), and the Xrn1 5′-3′ exonuclease (green). eIF4E is also present in P bodies, most likely bound to the 5′ cap on the silenced mRNA. The requirement of eIF4E-T for P-body assembly points to a central role for the cap plays a central role in the assembly process. The initial complex that contains silenced mRNA is submicroscopic, and the appearance of visible P bodies is thought to result from the aggregation of multiple silenced complexes. Little is known about the physical state of silenced mRNA in P bodies or what determines its fate. One possibility is that it is decapped and degraded (lower right). Alternatively, it might be stored in P bodies either with the cap intact or in a decapped state. We propose that the re-capping of decapped mRNA could facilitate its return to the translationally active pool. Given that poly(A) shortening precedes the translational silencing of most mRNAs, it seems likely that re-adenylation might also be a prerequisite for the re-activation of a stored or otherwise silenced mRNA.
References
-
- Lewis JD, Izaurralde E. The role of the cap structure in RNA processing and nuclear export. Eur J Biochem. 1997;247:461–469. - PubMed
-
- Gu M, Lima CD. Processing the message: structural insights into capping and decapping mRNA. Curr Opin Struct Biol. 2005;15:99–106. - PubMed
-
- Maquat LE. Nonsense-mediated mRNA decay: splicing, translation and mRNP dynamics. Nat Rev Mol Cell Biol. 2004;5:89–99. - PubMed
-
- Isken O, Maquat LE. Quality control of eukaryotic mRNA: safeguarding cells from abnormal mRNA function. Genes Dev. 2007;21:1833–1856. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
