

Establishment of cohesion at the pericentromere by the Ctf19 kinetochore subcomplex and the replication fork-associated factor, Csm3
- PMID: 19730685
- PMCID: PMC2727958
- DOI: 10.1371/journal.pgen.1000629
Establishment of cohesion at the pericentromere by the Ctf19 kinetochore subcomplex and the replication fork-associated factor, Csm3
Abstract
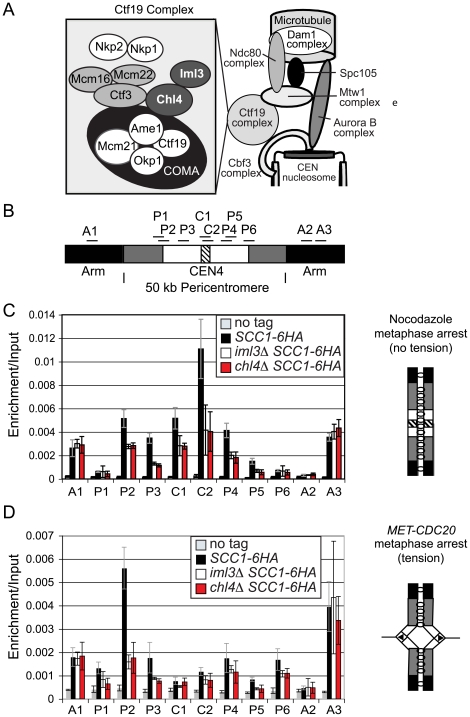
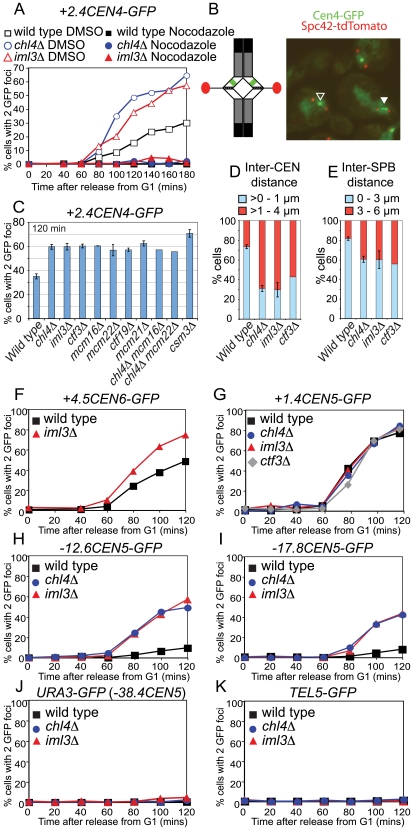
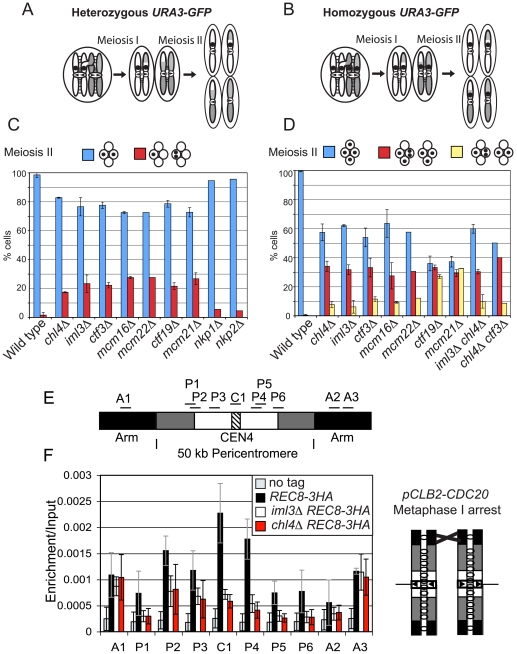
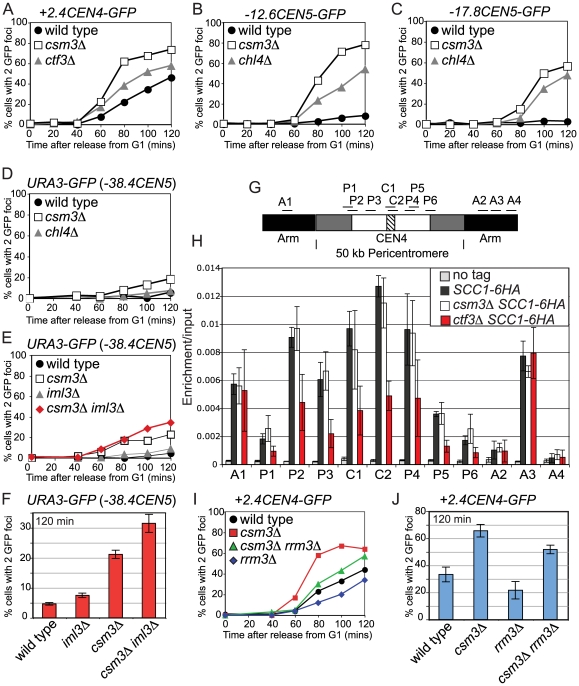
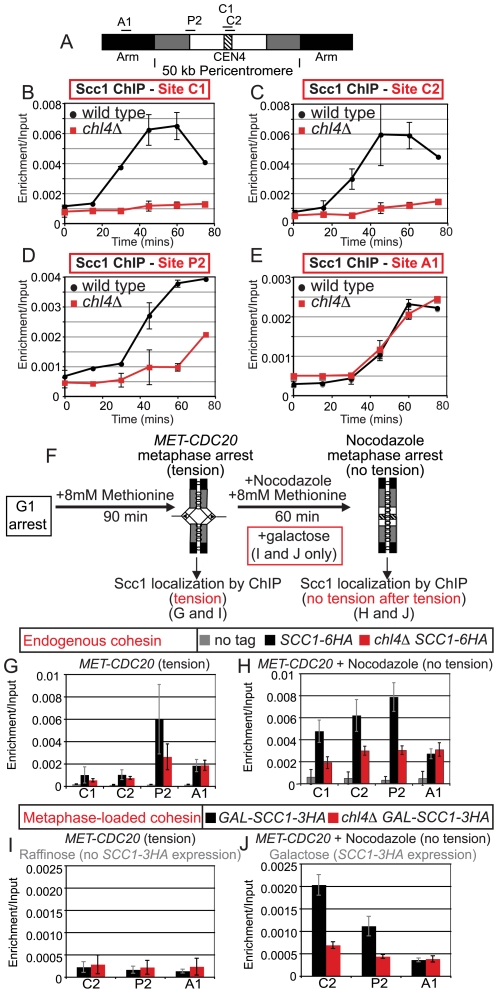
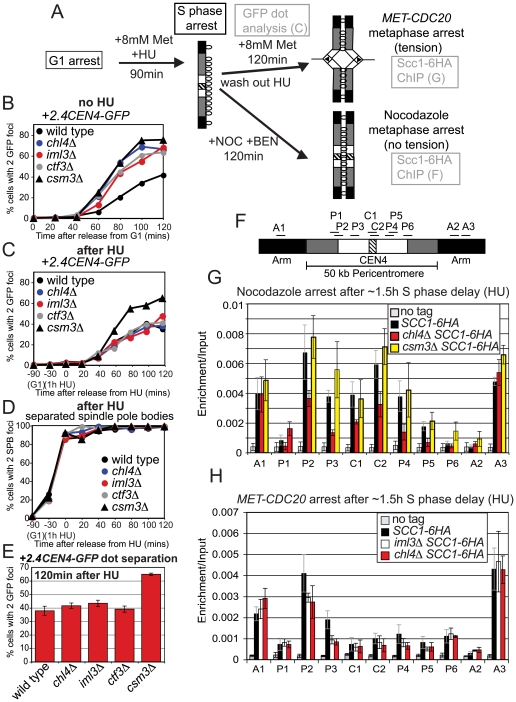
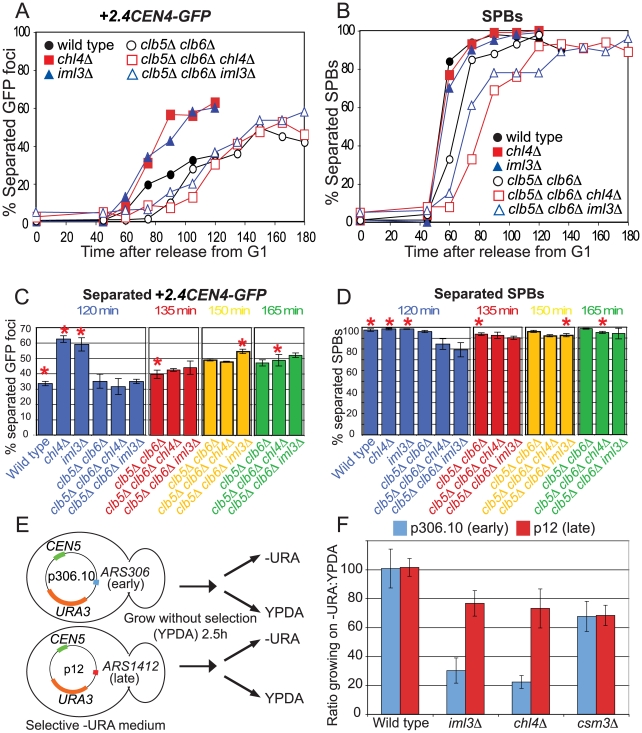
The cohesin complex holds sister chromatids together from the time of their duplication in S phase until their separation during mitosis. Although cohesin is found along the length of chromosomes, it is most abundant at the centromere and surrounding region, the pericentromere. We show here that the budding yeast Ctf19 kinetochore subcomplex and the replication fork-associated factor, Csm3, are both important mediators of pericentromeric cohesion, but they act through distinct mechanisms. We show that components of the Ctf19 complex direct the increased association of cohesin with the pericentromere. In contrast, Csm3 is dispensable for cohesin enrichment in the pericentromere but is essential in ensuring its functionality in holding sister centromeres together. Consistently, cells lacking Csm3 show additive cohesion defects in combination with mutants in the Ctf19 complex. Furthermore, delaying DNA replication rescues the cohesion defect observed in cells lacking Ctf19 complex components, but not Csm3. We propose that the Ctf19 complex ensures additional loading of cohesin at centromeres prior to passage of the replication fork, thereby ensuring its incorporation into functional linkages through a process requiring Csm3.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Nasmyth K. Disseminating the genome: joining, resolving, and separating sister chromatids during mitosis and meiosis. Annu Rev Genet. 2001;35:673–745. - PubMed
-
- Ciosk R, Shirayama M, Shevchenko A, Tanaka T, Toth A, et al. Cohesin's binding to chromosomes depends on a separate complex consisting of Scc2 and Scc4 proteins. Mol Cell. 2000;5:243–254. - PubMed
-
- Uhlmann F, Nasmyth K. Cohesion between sister chromatids must be established during DNA replication. Curr Biol. 1998;8:1095–1101. - PubMed
-
- Katou Y, Kanoh Y, Bando M, Noguchi H, Tanaka H, et al. S-phase checkpoint proteins Tof1 and Mrc1 form a stable replication-pausing complex. Nature. 2003;424:1078–1083. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
