

Molecular decay of the tooth gene Enamelin (ENAM) mirrors the loss of enamel in the fossil record of placental mammals
- PMID: 19730686
- PMCID: PMC2728479
- DOI: 10.1371/journal.pgen.1000634
Molecular decay of the tooth gene Enamelin (ENAM) mirrors the loss of enamel in the fossil record of placental mammals
Abstract
Vestigial structures occur at both the anatomical and molecular levels, but studies documenting the co-occurrence of morphological degeneration in the fossil record and molecular decay in the genome are rare. Here, we use morphology, the fossil record, and phylogenetics to predict the occurrence of "molecular fossils" of the enamelin (ENAM) gene in four different orders of placental mammals (Tubulidentata, Pholidota, Cetacea, Xenarthra) with toothless and/or enamelless taxa. Our results support the "molecular fossil" hypothesis and demonstrate the occurrence of frameshift mutations and/or stop codons in all toothless and enamelless taxa. We then use a novel method based on selection intensity estimates for codons (omega) to calculate the timing of iterated enamel loss in the fossil record of aardvarks and pangolins, and further show that the molecular evolutionary history of ENAM predicts the occurrence of enamel in basal representatives of Xenarthra (sloths, anteaters, armadillos) even though frameshift mutations are ubiquitous in ENAM sequences of living xenarthrans. The molecular decay of ENAM parallels the morphological degeneration of enamel in the fossil record of placental mammals and provides manifest evidence for the predictive power of Darwin's theory.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
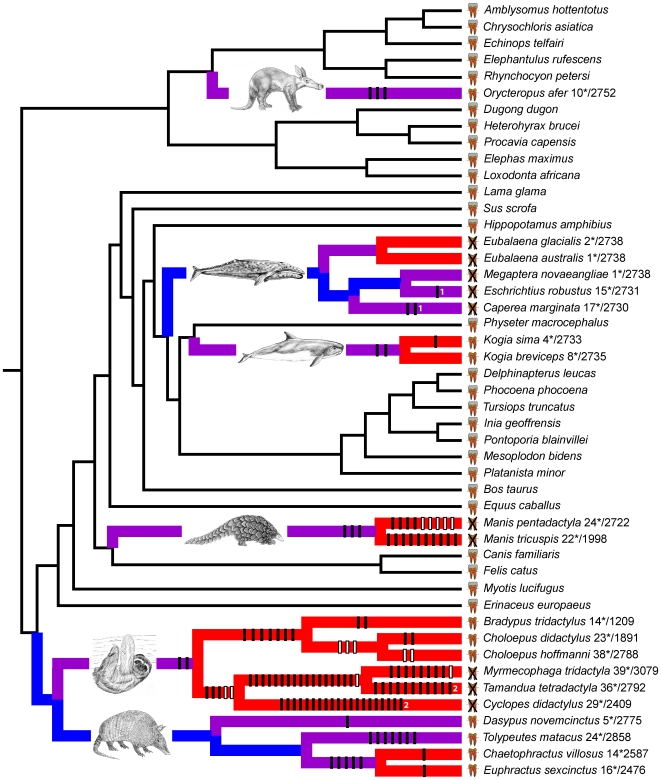
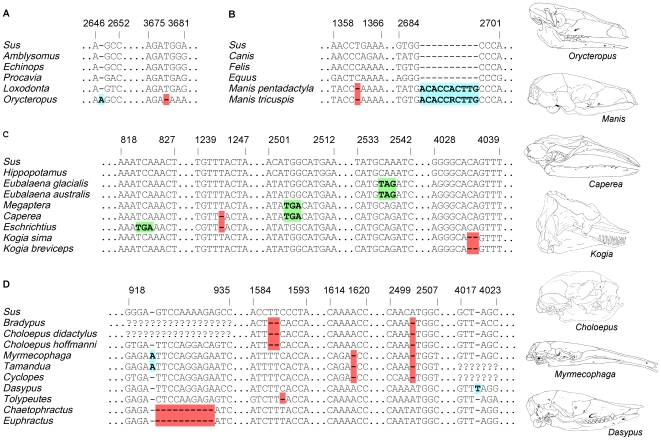
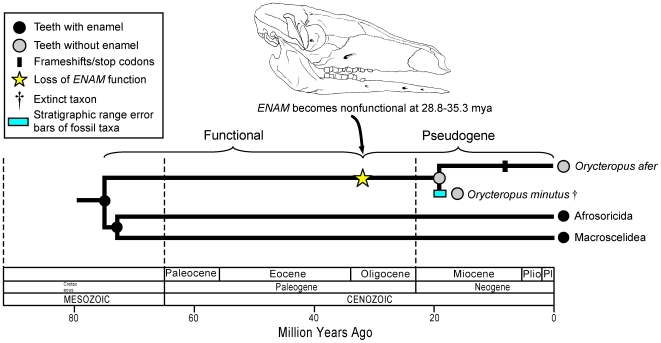
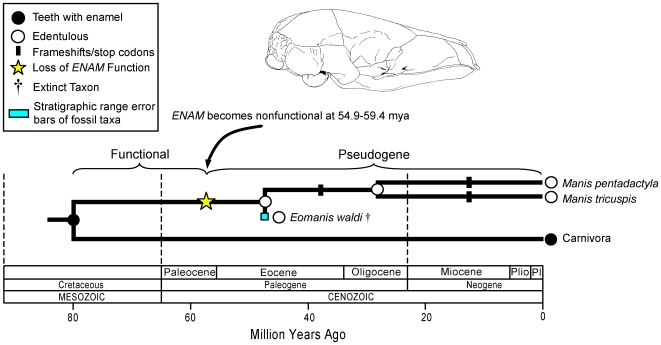
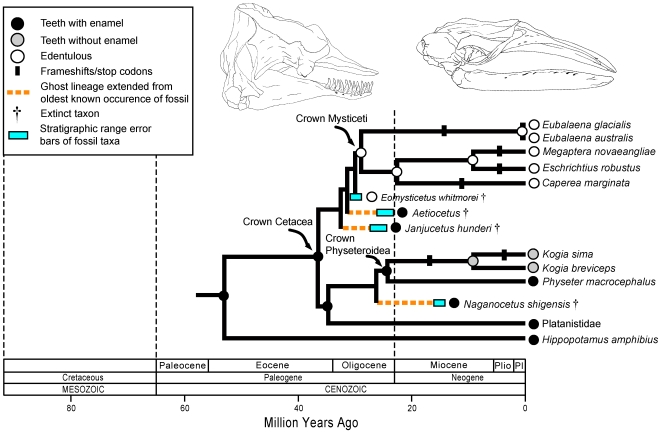
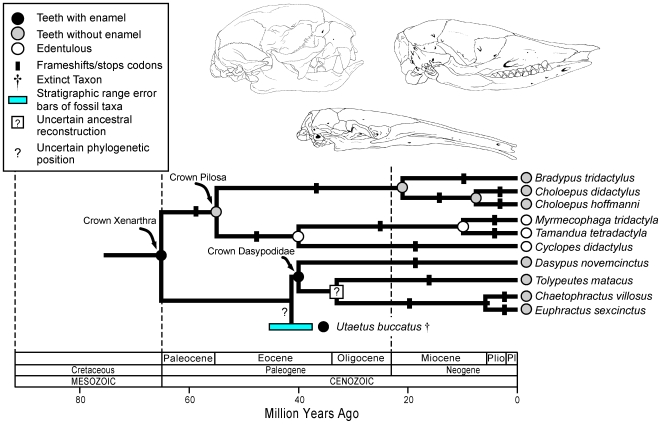
Comment in
-
Molecular tooth decay.PLoS Genet. 2009 Sep;5(9):e1000655. doi: 10.1371/journal.pgen.1000655. Epub 2009 Sep 18. PLoS Genet. 2009. PMID: 19763174 Free PMC article. No abstract available.
References
-
- Futuyma DJ. Sunderland: Sinauer; 1998. Evolutionary biology.
-
- Darwin C. London: John Murray; 1871. The descent of man, and selection in relation to sex.
-
- Bedjer L, Hall BK. Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss. Evol Dev. 2002;4:445–458. - PubMed
-
- Fong DW, Kane TC, Culver DC. Vestigialization and loss of nonfunctional characters. Annu Rev Ecol Syst. 1995;26:249–268.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
