

FKBP51 affects cancer cell response to chemotherapy by negatively regulating Akt
- PMID: 19732725
- PMCID: PMC2755578
- DOI: 10.1016/j.ccr.2009.07.016
FKBP51 affects cancer cell response to chemotherapy by negatively regulating Akt
Abstract
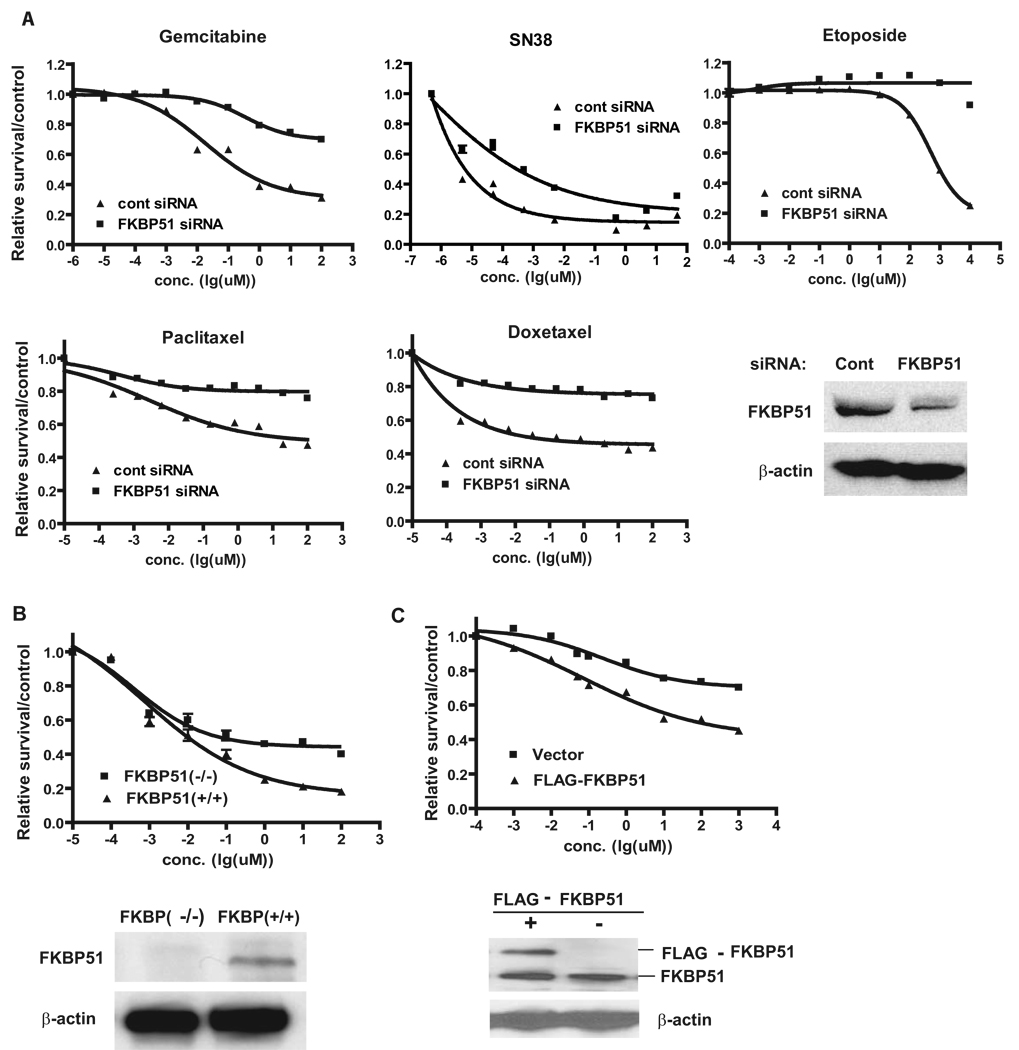
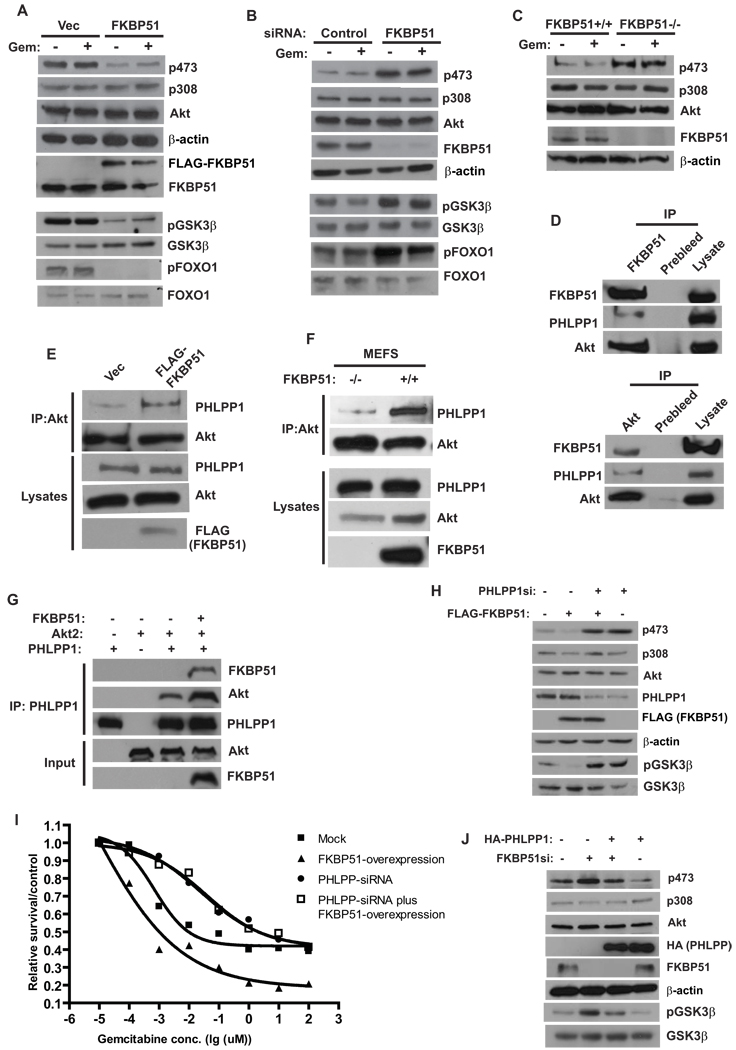
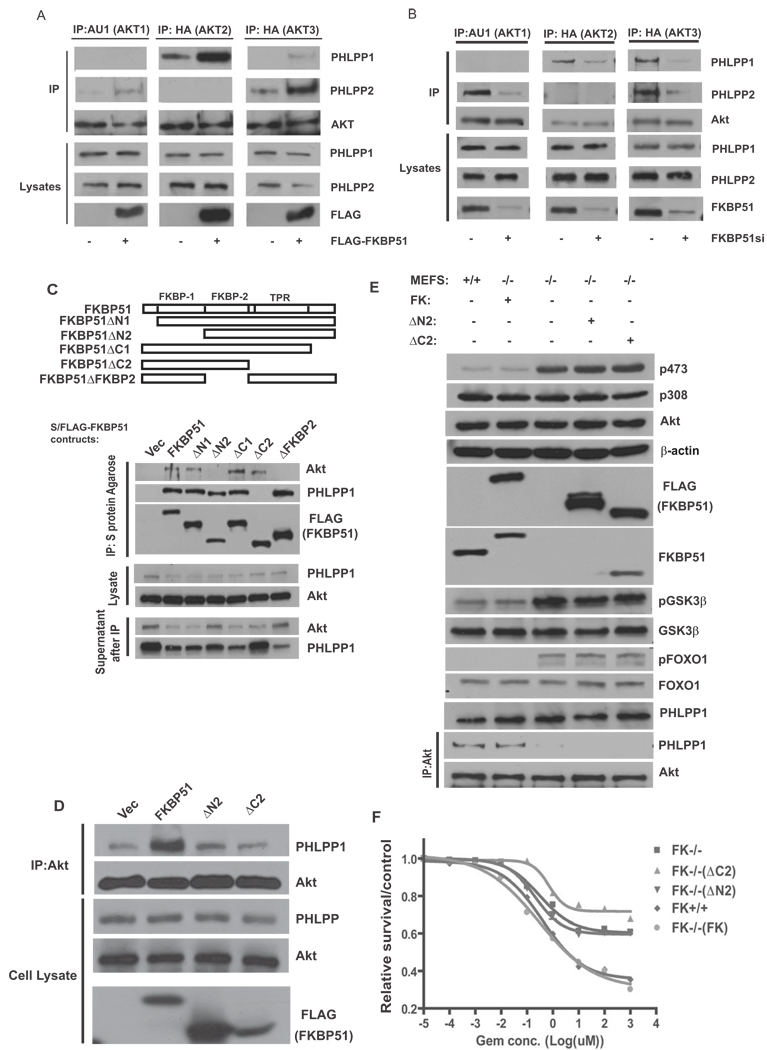
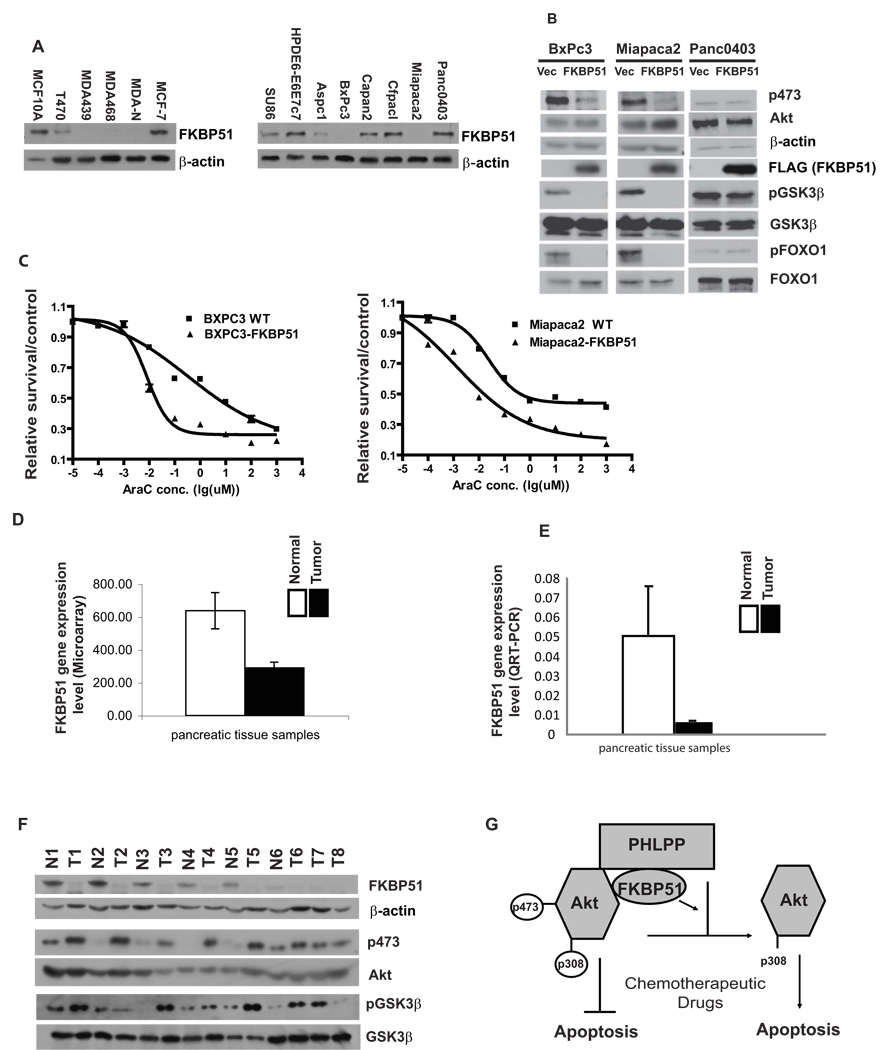
Akt is a central regulator of cell growth. Its activity can be negatively regulated by the phosphatase PHLPP that specifically dephosphorylates the hydrophobic motif of Akt (Ser473 in Akt1). However, how PHLPP is targeted to Akt is not clear. Here we show that FKBP51 (FK506-binding protein 51) acts as a scaffolding protein for Akt and PHLPP and promotes dephosphorylation of Akt. Furthermore, FKBP51 is downregulated in pancreatic cancer tissue samples and several cancer cell lines. Decreased FKBP51 expression in cancer cells results in hyperphosphorylation of Akt and decreased cell death following genotoxic stress. Overall, our findings identify FKBP51 as a negative regulator of the Akt pathway, with potentially important implications for cancer etiology and response to chemotherapy.
Figures
Comment in
-
A molecular link between AKT regulation and chemotherapeutic response.Cancer Cell. 2009 Sep 8;16(3):178-80. doi: 10.1016/j.ccr.2009.08.011. Cancer Cell. 2009. PMID: 19732717
References
-
- Alessi DR, James SR, Downes CP, Holmes AB, Gaffney PR, Reese CB, Cohen P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Balpha. Curr Biol. 1997;7:261–269. - PubMed
-
- Bai X, Ma D, Liu A, Shen X, Wang QJ, Liu Y, Jiang Y. Rheb activates mTOR by antagonizing its endogenous inhibitor, FKBP38. Science. 2007;318:977–980. - PubMed
-
- Barent RL, Nair SC, Carr DC, Ruan Y, Rimerman RA, Fulton J, Zhang Y, Smith DF. Analysis of FKBP51/FKBP52 chimeras and mutants for Hsp90 binding and association with progesterone receptor complexes. Mol Endocrinol. 1998;12:342–354. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
